
L'évolution nous a donné de nombreux animaux inhabituels. Certains d'entre eux sont si inhabituels que leurs découvreurs ont été appelés des escrocs (comme ce fut le cas avec les ornithorynques en 1797). Mais non seulement la faune peut se vanter d'espèces non standard, mais parmi les représentants de la flore, il y a aussi des corbeaux blancs, à savoir des plantes qui ne veulent pas s'asseoir sur un régime ensoleillé, mais préfèrent les insectes juteux et autres petits animaux comme plat du jour. Plus tôt, nous avons déjà pris connaissance d'une étude dans laquelle des scientifiques ont examiné le piège à mouches de Vénus et ses épines marginales.... Aujourd'hui, nous devons découvrir à quel point ces épines sont sensibles. Des scientifiques de l'Université de Zurich (Suisse) ont mené une série d'expériences pratiques dont le but était de mesurer la vitesse de réaction du piège à mouches Vénus au contact d'une force particulière. Comment exactement les scientifiques ont-ils chatouillé le moucherolle, à quelle vitesse a-t-il réagi et comment ces résultats expliquent-ils les préférences gastronomiques de la plante prédatrice? Nous trouverons des réponses à ces questions dans le rapport des scientifiques. Aller.
Base de recherche
Afin de ne pas étirer ce récit, la description du piège à mouches Vénus (qui était déjà utilisé dans l'article précédent) est cachée sous le spoiler:
Qu'est-ce qu'un flycatcher?
, . , . — , . , «» (, ) ( , ). 630 , , .
: . ( ), — , .

. , — ( 300 000 1 ) , . , , . , . , .
, «».
, . : , , . , , . , , , . -, . .

: , - .
, -, . . , . , . , . , ? , . 2 20 . , , . , , , , .
. , . , , . , ( ), , . . , , . — , . , . , 2-3 .
. , , . , , . , .
True Facts: Carnivorous Plants ( : ). .
, -. , , .
, , , , « ». , , «». , , , , . , . , .
: . ( ), — , .
. , — ( 300 000 1 ) , . , , . , . , .
, «».
, . : , , . , , . , , , . -, . .
: , - .
, -, . . , . , . , . , ? , . 2 20 . , , . , , , , .
. , . , , . , ( ), , . . , , . — , . , . , 2-3 .
. , , . , , . , .
True Facts: Carnivorous Plants ( : ). .
, -. , , .
, , , , « ». , , «». , , , , . , . , .
L'idée principale du mécanisme de déclenchement du piège du piège à mouches Vénus est que la future proie doit faire deux touchers sur les poils sensibles de la plante dans les 30 secondes. Chaque contact produit un potentiel d'action * nécessaire pour fermer le piège. Les scientifiques le croient depuis 200 ans, mais la théorie est-elle cohérente avec la pratique?
Au fil des années de recherche, les scientifiques ont déduit un scénario d'événements qui devraient (en théorie) se dérouler lors d'une chasse au moucherolle:
- une plante affamée attire les insectes en sécrétant des composés volatils;
- l'insecte peu averti explore la plante (en espérant de la nourriture gratuite), au cours de laquelle il touche l'un des six poils sensibles, provoquant ainsi le potentiel d'action (PA du potentiel d'action);
- 30 , «It’s a trap!», , ;
- (, ), (C12H18O3) .
En conséquence, les «pétales» précédemment ouverts du moucherolle se transforment en une sorte d'estomac, où les proies infructueuses seront digérées.
Dans ce travail, les scientifiques ont décidé d'examiner en détail le processus de conversion de la stimulation mécanique des cheveux sensibles en un signal électrique. Nous savons avec certitude que toucher des poils sensibles ouvre des canaux ioniques mécanosensibles, mais nous n'avons pas encore été en mesure d'identifier ces canaux.
Alors que ces canaux putatifs sont ouverts, le potentiel récepteur * (RP du potentiel récepteur ) augmente, et si la déflexion des cheveux est suffisamment grande, RP atteint un seuil au-dessus duquel AP se manifeste.
* — , .
* — , .
* — .
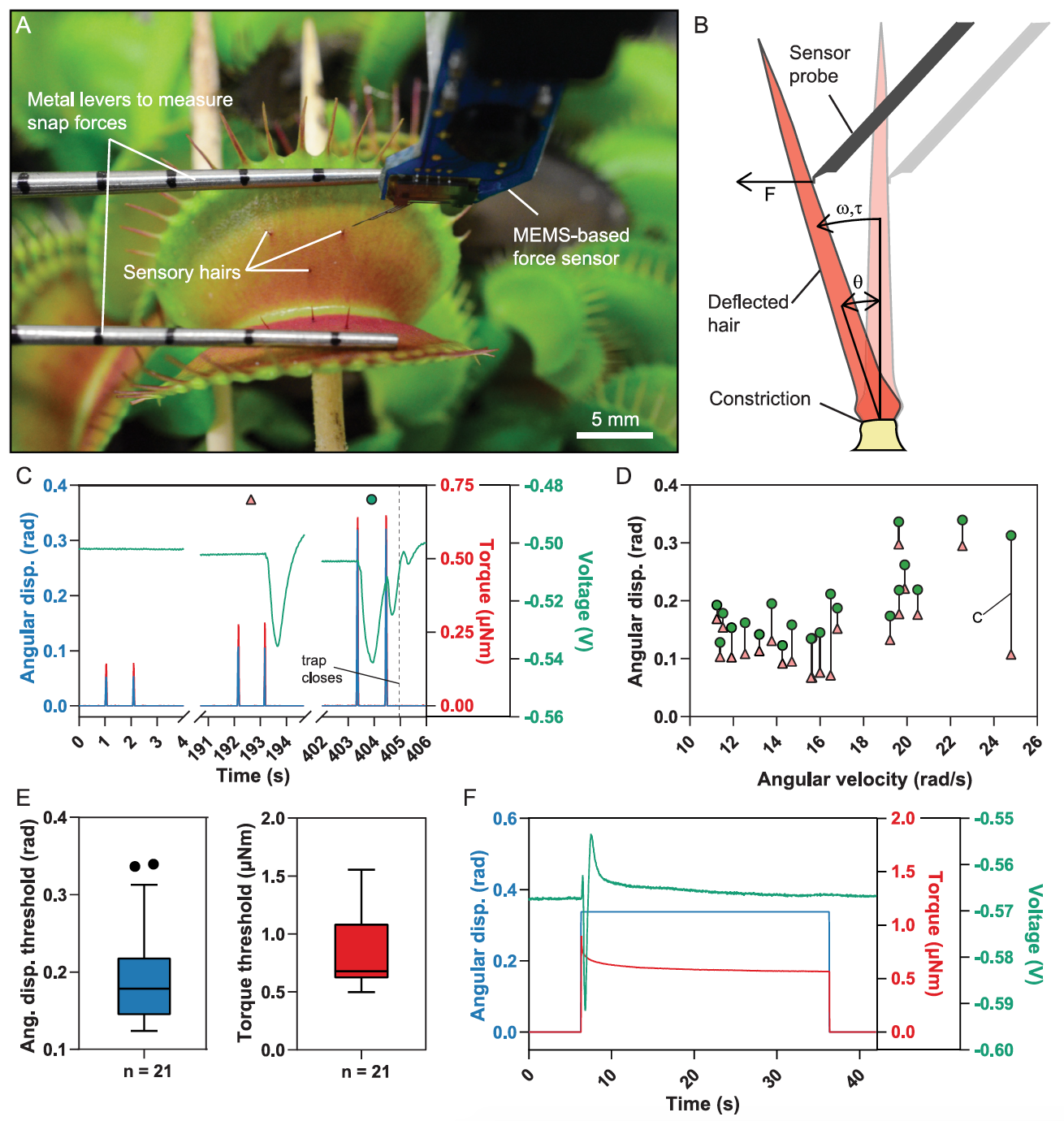
Image # 1
Il y a eu des tentatives antérieures de lier les stimuli mécaniques et la génération de potentiel d'action, mais il n'y avait pas d'instruments appropriés pour des mesures précises. Dans le cas de l'étude que nous examinons aujourd'hui, les scientifiques ont utilisé un capteur de force à base de MEMS * monté sur un système microrobotique pour contrôler précisément la vitesse et l'amplitude de la déflexion, et mesurer simultanément la force appliquée ( 1A et 1B ).
Les MEMS * (systèmes microélectromécaniques) sont des dispositifs qui combinent des composants microélectroniques et micromécaniques.
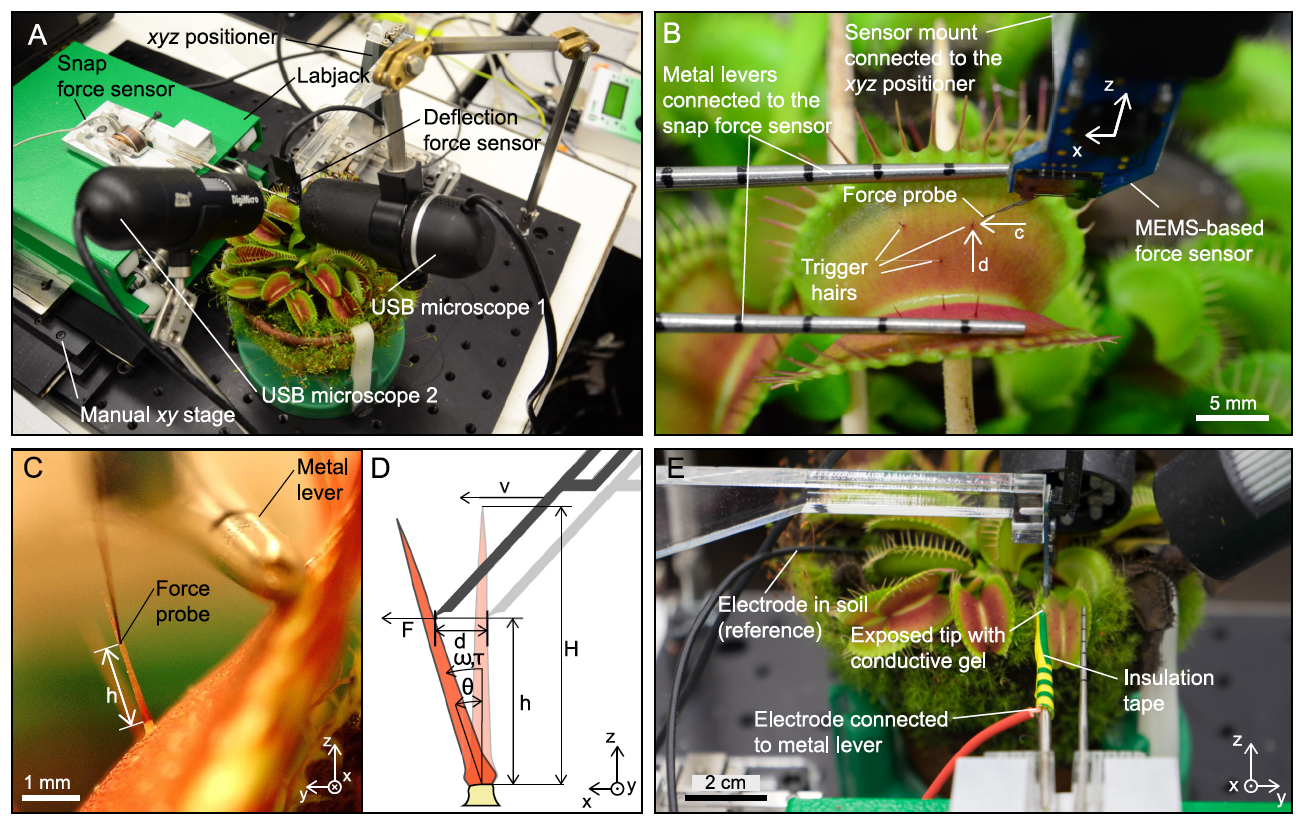
Image # 2
De cette manière, les scientifiques ont pu déterminer avec précision la gamme de paramètres dans lesquels la déflexion des cheveux conduit à la fermeture du piège, tandis que le deuxième capteur de force mesurait la force instantanée générée (image ci-dessus).
Résultats de recherche
Au cours des expériences, les scientifiques ont pris en compte le fait que les araignées, les mouches et les fourmis (proie commune d'un moucherolle) provoqueraient une déformation assez rapide des poils sensibles. Par conséquent, le système microrobotique a été déployé à pleine vitesse pour imiter ces stimuli. Il en résulte une vitesse angulaire initiale élevée allant de 10 à 20 rad / s.
Des études antérieures ont montré que les fourmis dévient les poils sensibles à une vitesse angulaire de 0,25 à 7,8 rad / s, mais plus lentement que les mouches. Par conséquent, les paramètres utilisés sont plus élevés que les vrais, ce qui est bien adapté pour évaluer les capacités du piège à mouches Venus.
À des vitesses aussi élevées, la durée de la déflexion est significativement plus courte que les autres facteurs dépendants du temps impliqués, tels que la dégradation de la RP et la relaxation sensorielle des cheveux (1F ).
La prise en compte de l'écart angulaire plutôt que linéaire a permis de corriger les différences de hauteur de contact de la sonde sensorielle par rapport au lieu de rétrécissement des cheveux sensoriels, ainsi que les différences concernant les différentes géométries des cheveux sensoriels ( 1B , 2C et 2D ). Par conséquent, une déflexion peut être approchée par une augmentation discrète du décalage angulaire, et le déclenchement AP dépend principalement de l'amplitude du décalage angulaire.
Une déflexion a été définie comme une combinaison de déplacement angulaire avant-arrière, similaire à ce qui se produit lorsqu'une proie touche un poil. Chaque mesure consistait en deux déflexions successives avec un intervalle de 1 s entre elles jusqu'à un déplacement angulaire donné θ. Si le piège ne se fermait pas, une phase d'attente (2 minutes) suivait pour s'assurer que le RP était complètement réinitialisé. L'attente d'exactement deux minutes, et pas plus / moins, est due au fait qu'une série d'expériences antérieures a démontré qu'à des températures inférieures à 30 ° C, deux déflexions des cheveux en 30 à 40 s sont nécessaires pour fermer rapidement et complètement le piège.
Après la phase d'attente, la procédure a été répétée avec des déplacements angulaires croissants jusqu'à ce que le mécanisme de fermeture du piège soit déclenché ( 1C et 1D). Cela s'est produit lorsque le seuil de déplacement moyen θ = 0,18 rad ou le seuil de couple moyen τ = 0,8 μN · m (n = 21) a été atteint.
Les scientifiques notent que la fermeture du piège n'a jamais été enregistrée si θ était inférieur à 0,12 rad et τ était inférieur à 0,50 μN · m ( 1E ). Par conséquent, ces indicateurs sont la limite inférieure de l'écart angulaire nécessaire pour déclencher la fermeture du piège dans les conditions de cette expérience. Il a également été possible de déterminer que pour que le piège fonctionne, l'insecte doit appliquer une force (F) de 0,5 mN près de l'extrémité des poils ou de 5 mN près de la base.
Les mesures AP ont fourni un lien entre la déviation sensorielle des poils et la fermeture du piège. Lorsque deux écarts consécutifs étaient bien en dessous du seuil de biais (θ << 0,12 rad), aucun PA n'a été observé. Pour des amplitudes de déflexion proches du seuil de déplacement (θ <0,12 rad), un seul PA a été détecté après la deuxième déflexion.
Cela indiquait que les deux déflexions des cheveux contribuaient à la RP et que le seuil d'induction AP n'était atteint qu'à la deuxième déflexion. Comme prévu, un seul AP n'était pas suffisant pour déclencher le piège.
L'hypothèse selon laquelle chaque contact déclenche AP n'est valable que si la déviation des poils sensoriels dépasse le seuil de déplacement. Si tel est le cas, deux points d'accès (un pour chaque déflexion des cheveux) ont été générés et ont provoqué la fermeture du piège ( 1C ).
Double déflexion des poils sensibles résultant en deux AP et fermeture du piège.
Ces résultats suggèrent qu'une déflexion rapide des poils sensibles augmente la RP à un certain niveau qui dépend de l'amplitude de la déflexion angulaire. Les RP peuvent s'empiler et invoquer des PA après plusieurs déflexions s'ils sont en dessous du seuil de déviation. Cependant, un PA par contact n'est généré que si la déflexion des poils sensoriels est supérieure au seuil de déflexion.
En d'autres termes, le potentiel d'action ne sera généré que lorsque les cheveux sensibles auront été suffisamment «perturbés». Mais un potentiel d'action unique ne suffit pas pour déclencher la fermeture du piège. Cela correspond bien à la théorie selon laquelle les flycatchers ne gaspilleront pas d'énergie pour fermer le piège chaque fois que quelque chose ou quelqu'un touche leurs cheveux. Sinon, il peut y avoir de nombreux pièges inactifs sans retour d'énergie (c'est-à-dire nutritionnel) (la proie a réussi à s'échapper, la proie était trop petite ou ce n'était pas du tout une proie, mais des ordures).
Étant donné que l'augmentation de RP avec de multiples écarts est additive, il serait logique de supposer qu'un déplacement soutenu (à long terme) d'un cheveu peut avoir un effet similaire.
Pour tester cette hypothèse, les scientifiques ont rejeté les cheveux sensibles du moucherolle au-delà du seuil de déplacement angulaire et l'ont verrouillé dans cette position pendant 30 secondes ( 1F ). Cependant, cela n'a pas déclenché les pièges.
Le déplacement initial a provoqué un seul AP, après quoi la tension est rapidement revenue à la ligne de base, bien que les poils soient restés déviés.
Si le biais soutenu devait contribuer au RP, il devrait rester au-dessus du seuil, auquel cas une série AP était attendue. Des analyses supplémentaires ont montré que le désalignement angulaire joue un rôle important dans la RP, mais la déviation statique des cheveux n'y contribue en aucune manière.
Il est curieux que dans les expériences précédentes, la situation était différente: un seul déplacement des poils a conduit à la fermeture du piège. Cependant, il ne s'agissait pas d'un déplacement unique intégral, mais de nombreux petits déplacements, puisque les oscillations qui s'accompagnent d'une déflexion manuelle d'un cheveu sont probablement supérieures au seuil de déplacement angulaire.
L'étape suivante de l'étude a consisté à créer un modèle d' accumulation de charge électromécanique (ECB ), qui prédit de manière surprenante la fermeture des pièges à partir d'un seul toucher.
Sur la base des données obtenues lors des expériences, les scientifiques ont développé un modèle simple pour étudier les limites de déplacement angulaire et de vitesse à l'intérieur desquelles les pièges vont réagir.
Dans le modèle ECB, la déflexion mécanique conduit à une accumulation de charge RP en fonction de la vitesse angulaire ω et du déplacement θ, tandis que les charges sont dissipées en continu. Si les charges accumulées dépassent un certain seuil Q th RP , un AP se produit. De plus, une période réfractaire t RP a été mise en œuvre , représentant l'intervalle de temps nécessaire pour qu'un autre PA soit activé.
En conséquence, le modèle a prédit que si les écarts sont trop rapides et / ou trop petits, alors plus de deux de ces écarts peuvent être nécessaires pour provoquer la fermeture du piège (zone rouge à 3A ).

Image # 3
Cela est dû au fait qu'un écart ne suffit pas pour activer AP ( 3B), comme le montrent des expériences pratiques ( 1C , au centre).
De même, le modèle a montré que de très faibles vitesses angulaires (ω <0,04 rad / s) ne peuvent pas remplir RP.
Mais le résultat le plus inattendu de la simulation a été la prédiction de la plage de vitesses angulaires intermédiaires (0,04 rad / s <ω <10 rad / s), à laquelle une déflexion suffit pour activer deux ou plusieurs AP, nécessaires pour fermer le piège ( 3B ).
Cependant, une telle prédiction contredit l'idée du fonctionnement du mécanisme de piège à mouches Vénus. Par conséquent, les scientifiques ont décidé de vérifier si cette situation de modèle peut être réalisée dans la pratique.
Une seule déviation des poils sensibles entraînant la fermeture du piège.
De manière surprenante, la fermeture du piège s'est effectivement produite avec une seule déflexion des poils si les vitesses angulaires étaient suffisamment faibles.
Pour réduire la plage de valeurs dans laquelle cela se produit, les scientifiques ont dévié à plusieurs reprises les mêmes poils sensoriels à différentes vitesses angulaires jusqu'à ce que le piège se ferme. Il y avait un intervalle de 2 minutes entre deux déflexions successives jusqu'à ce que le piège se rétablisse et que le RP soit épuisé.
La limite inférieure de la vitesse angulaire ω nécessaire pour fermer le piège à une déviation a été déterminée en augmentant progressivement la vitesse angulaire après chaque déviation (n = 17). La vitesse initiale était inférieure à 0,009 rad / s, car le piège n'a jamais été fermé à cette vitesse.
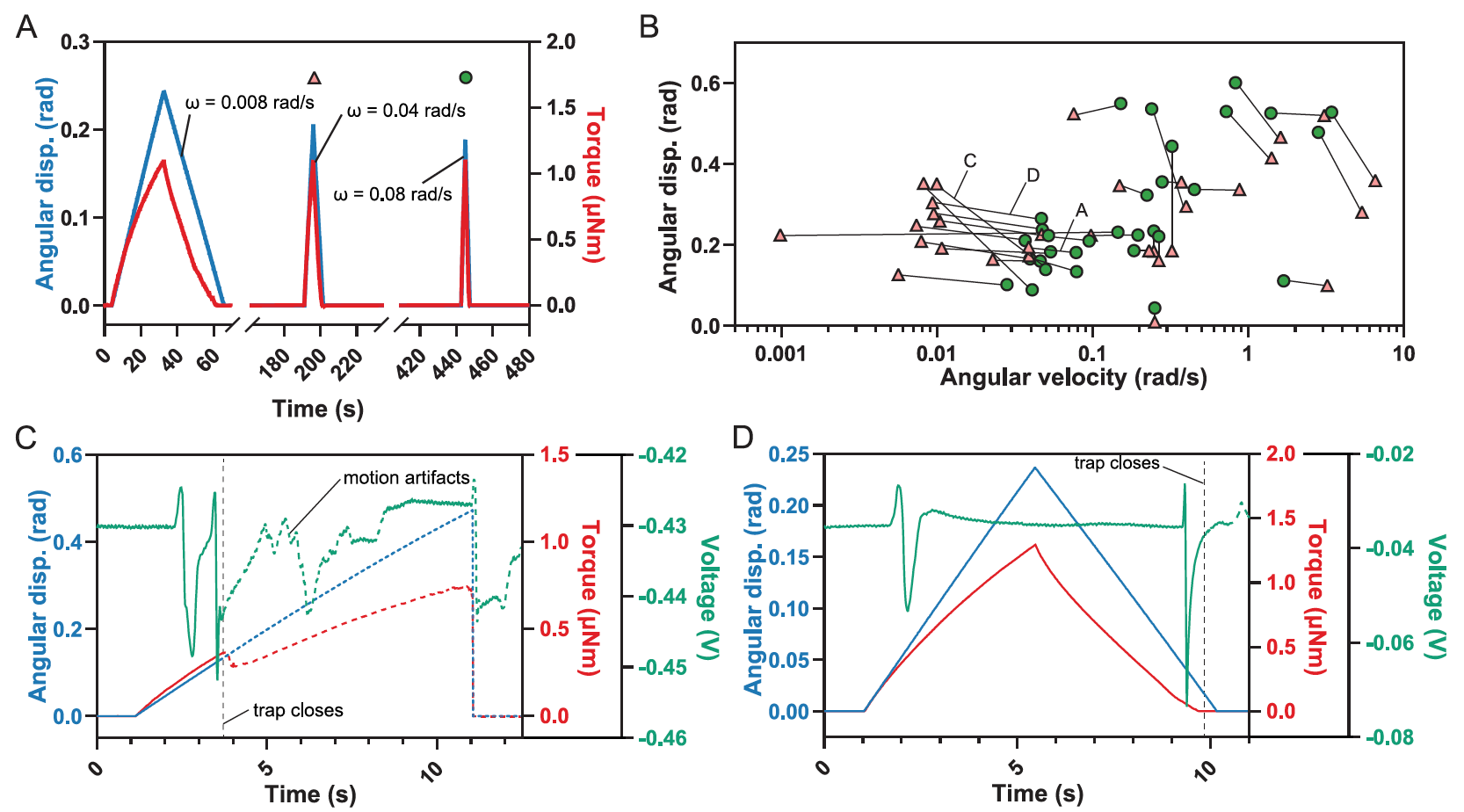
Image # 4
Des déflexions de poils uniques subséquentes ont été effectuées avec une augmentation graduelle de la vitesse jusqu'à ce que le piège soit déclenché ( 4A ).
La limite supérieure a été déterminée de manière similaire, en commençant par une vitesse de ω> 3 rad / s suivie d'une décroissance par étapes (n = 9).
De plus, une autre série d'expériences avec une déflexion (n = 5) a été réalisée, dans laquelle la vitesse de la sonde de force a été maintenue constante, ce qui a conduit à une vitesse angulaire intermédiaire entre 0,2 et 0,4 rad / s, tandis que le déplacement angulaire θ a augmenté progressivement pendant le temps des déflexions ultérieures pour obtenir la borne inférieure θ nécessaire au fonctionnement du piège dans les conditions d'une seule déflexion des poils.
Toutes les déviations simples menant à la fermeture du piège, ainsi que les stimuli antérieurs pour lesquels aucune fermeture de piège ne s'est produite, ont identifié la région où une seule déviation provoque la fermeture ( 4B ).
Une seule déviation peut provoquer la fermeture du piège à des vitesses angulaires de déviation intermédiaires (0,03 rad / s ≤ ω ≤ 4 rad / s). Mais la déflexion seule à des vitesses inférieures ou supérieures ne suffira pas.
Des expériences, où la vitesse de déflexion des cheveux était plutôt faible, ont montré que le piège se déclenchait à la fois lorsque le capteur s'approchait de la base des cheveux et lorsque le capteur s'en éloignait.
Déflexion unique des poils sensibles résultant en deux AP lors de la flexion initiale, entraînant la fermeture du piège.
Lorsque le piège a été refermé lors de l'approche de la sonde, deux potentiels d'action successifs ( 4C ) ont été observés lors de la déflexion des cheveux . Lorsque le piège a été refermé lors du retrait de la sonde, un PA est apparu au moment de la déflexion du poil, et le second après le retrait de la sonde, lorsque le poil est revenu à sa position d'origine ( 4D ). Dans les deux cas, le deuxième RA a conduit à la fermeture immédiate du piège.
En conclusion, les scientifiques ont décidé de mesurer la force de fermeture du piège ( 5A ).
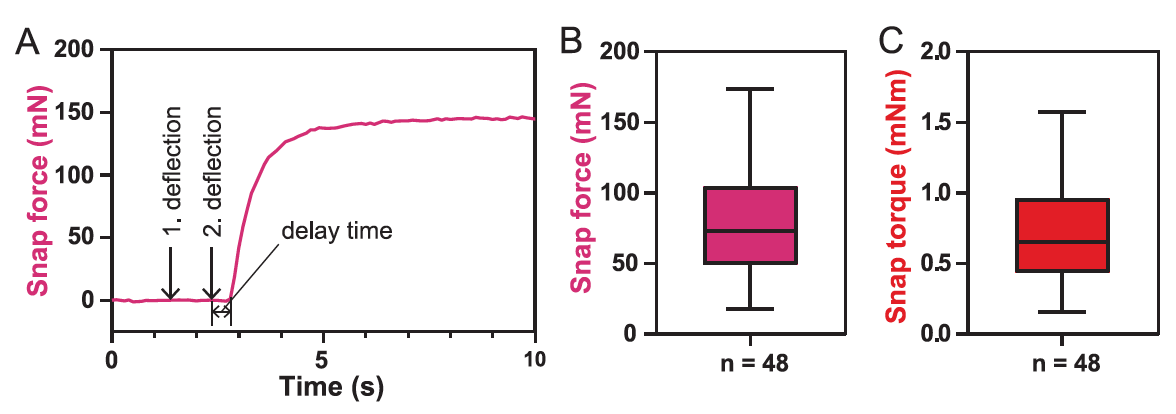
Image n ° 5
Pour le piège 48, la valeur moyenne de la force de préhension F close était de 73 mN ( 5V), et la plage de valeurs était de 18 à 174 mN.
Ces valeurs sont inférieures à celles publiées précédemment (140-150 mN). Il y a une explication à cela: dans ce cas, la force a été mesurée au début de la fermeture du piège, et dans les observations précédentes, la force agissant sur les bords des feuillets du piège à feuilles à la fin de la fermeture a été mesurée.
Étant donné que la force mesurée dépend fortement de la position du transducteur ainsi que de l'orientation et de la taille de la feuille, le couple de fermeture τ se ferme autour du milieu de la feuille avec une valeur moyenne de 0,65 mN · m est la meilleure valeur pour décrire la force de fermeture du piège ( 5C ). Le temps de retard, c'est-à-dire le temps entre le stimulus mécanique et le début de la fermeture du piège, était de 0,6 ± 0,3 s.
Pour une connaissance plus détaillée des nuances de l'étude, je vous recommande de regarderrapport des scientifiques .
Épilogue
Le piège à mouches de Vénus ne court pas comme un guépard, n'a pas de poison mortel comme un cobra et n'entend certainement pas sa proie comme un hibou. Mais cette plante est en effet l'un des meilleurs carnivores de la planète, car son mécanisme de chasse fonctionne comme une horloge.
Dans ce travail, les scientifiques ont pu déterminer quels devraient être ces ou ces paramètres pour que le piège fonctionne. Comme il s'est avéré que, dans la plupart des cas, pour déclencher le piège, la victime doit faire deux erreurs consécutives: d'abord, toucher les poils sensoriels du piège à mouches; la seconde est de recommencer.
La déviation répétée du cheveu par rapport à sa position initiale conduit à la génération d'un second potentiel d'action, qui déclenche le processus de fermeture du piège. Néanmoins, la simulation a montré que dans certaines conditions, une seule touche peut suffire, mais en réalité, cela est très peu probable.
En d'autres termes, le piège à mouches Vénus peut-il servir de chasseur de moustiques? Non, car ces sangsues sont très petites et trop agiles. Mais le moucherolle ne regrette ni les mouches ni les escargots.
Merci pour votre attention, restez curieux et passez un excellent week-end, les gars! :)
Un peu de publicité
Merci de rester avec nous. Aimez-vous nos articles? Vous voulez voir du contenu plus intéressant? Soutenez-nous en passant une commande ou en recommandant à des amis, Cloud VPS pour les développeurs à partir de 4,99 $ , un analogue unique des serveurs d'entrée de gamme que nous avons inventé pour vous: The Whole Truth About VPS (KVM) E5-2697 v3 (6 Cores) 10 Go DDR4 480 Go SSD 1Gbps à partir de 19 $ ou comment diviser correctement le serveur? (options disponibles avec RAID1 et RAID10, jusqu'à 24 cœurs et jusqu'à 40 Go de DDR4).
Le Dell R730xd 2x est-il moins cher dans le centre de données Equinix Tier IV à Amsterdam? Seulement, nous avons 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2,6 GHz 14C 64 Go DDR4 4x960 Go SSD 1 Gbps 100 TV à partir de 199 $ aux Pays-Bas!Dell R420 - 2 x E5-2430 2,2 GHz 6C 128 Go DDR3 2 x 960 Go SSD 1 Gbps 100 To - À partir de 99 $! Lisez à propos de Comment construire l'infrastructure de bldg. classe avec les serveurs Dell R730xd E5-2650 v4 au coût de 9000 euros pour un centime?