
À quoi associez-vous l'été? Pour certains, c'est une période de vacances tant attendues, pour quelqu'un - des vacances, et pour quelqu'un - chaleur, étouffement et inconfort. Si nous considérons l'été du point de vue de la gastronomie, c'est la période des légumes, des fruits et des baies, que nous aimons non seulement pour leur goût et leurs bienfaits, mais aussi pour leur apparence. Comme on le sait depuis le cours initial de biologie, les fruits de nombreuses plantes ont certaines propriétés dont le but est d'attirer un potentiel gourmet. C'est un élément important des tactiques d'expansion de la zone de croissance. La grande majorité des fruits ont une couleur vive et juteuse, indiquant leur bonté. La principale source de telle ou telle couleur dans les baies est les pigments dans la peau, mais ce n'est pas la seule technique de teinture. Des scientifiques de l'Université de Bristol ont découvert que le laurier viburnum ( Viburnum tinus) utilise des nanostructures lipidiques dans les parois cellulaires pour colorer leurs baies, une variante jusqu'alors inconnue de la coloration structurelle. Qu'y a-t-il de si inhabituel dans ces nanostructures lipidiques qui donnent aux baies une couleur bleu foncé, et quelle est l'application pratique de cette découverte? Le rapport des scientifiques éclairera ces questions Aller.
Résultats de recherche
Le protagoniste de cette œuvre est le laurier viburnum ( Viburnum tinus / viburnum tinus) - un buisson ou un arbre à feuilles persistantes atteignant 6 mètres de hauteur, poussant dans la région méditerranéenne.

Viburnum tinus pendant la floraison.
Viburnum tinus porte des fruits plusieurs fois au cours de l'année avec des baies bleu foncé, fournissant de la nourriture à de nombreuses espèces d'oiseaux, y compris la paruline à tête noire (Sylvia atricapilla) et le merle (Erithacus rubecula). Comme pour toute autre plante à baies, les oiseaux pour V. tinus viburnum sont la principale méthode de propagation des graines vers de nouveaux territoires.

Sylvia atricapilla (à gauche) et Erithacus rubecula (à droite).
À première vue, il n'y a rien de spécial à propos de cette plante. Un bel arbuste à feuilles persistantes qui ravit les esthètes parmi les gens et les gourmets parmi les oiseaux. Cependant, un examen détaillé des baies suggère le contraire. Alors que d'autres plantes utilisent différents composés chimiques pour colorer leurs fruits, le viburnum tinus utilise une coloration structurelle. Bien que l'on pensait auparavant que la couleur bleue des baies de V. tinus était causée par la présence de pigments anthocyaniques dans leur peau, ce n'est naturellement pas vrai.
Les couleurs structurelles sont assez courantes chez la faune: ailes de papillon, coquilles de coléoptère, plumes de paon, etc. La couleur dans leur boîtier est formée en raison des caractéristiques structurelles nanométriques de la surface, provoquant des interférences avec la lumière visible.

Exemples de couleurs structurelles dans la nature: A- hibiscus triangulaire (Hibiscus trionum); B - coléoptère tamamusi (Chrysochroa fulgidissima); C - papillon de l'espèce Morpho rhetenor; D - moustique commun (Culex pipiens); E - souris de mer (Aphrodita aculeata); F - coléoptère de l'espèce Pachyrhynchus argus; G - un papillon de l'espèce Parides sesostris.
La particularité de Viburnum Tinus, qui a attiré l'attention des scientifiques, est qu'il démontre non seulement un nouveau mécanisme de coloration structurelle, mais est également l'une des rares plantes capables de cela.
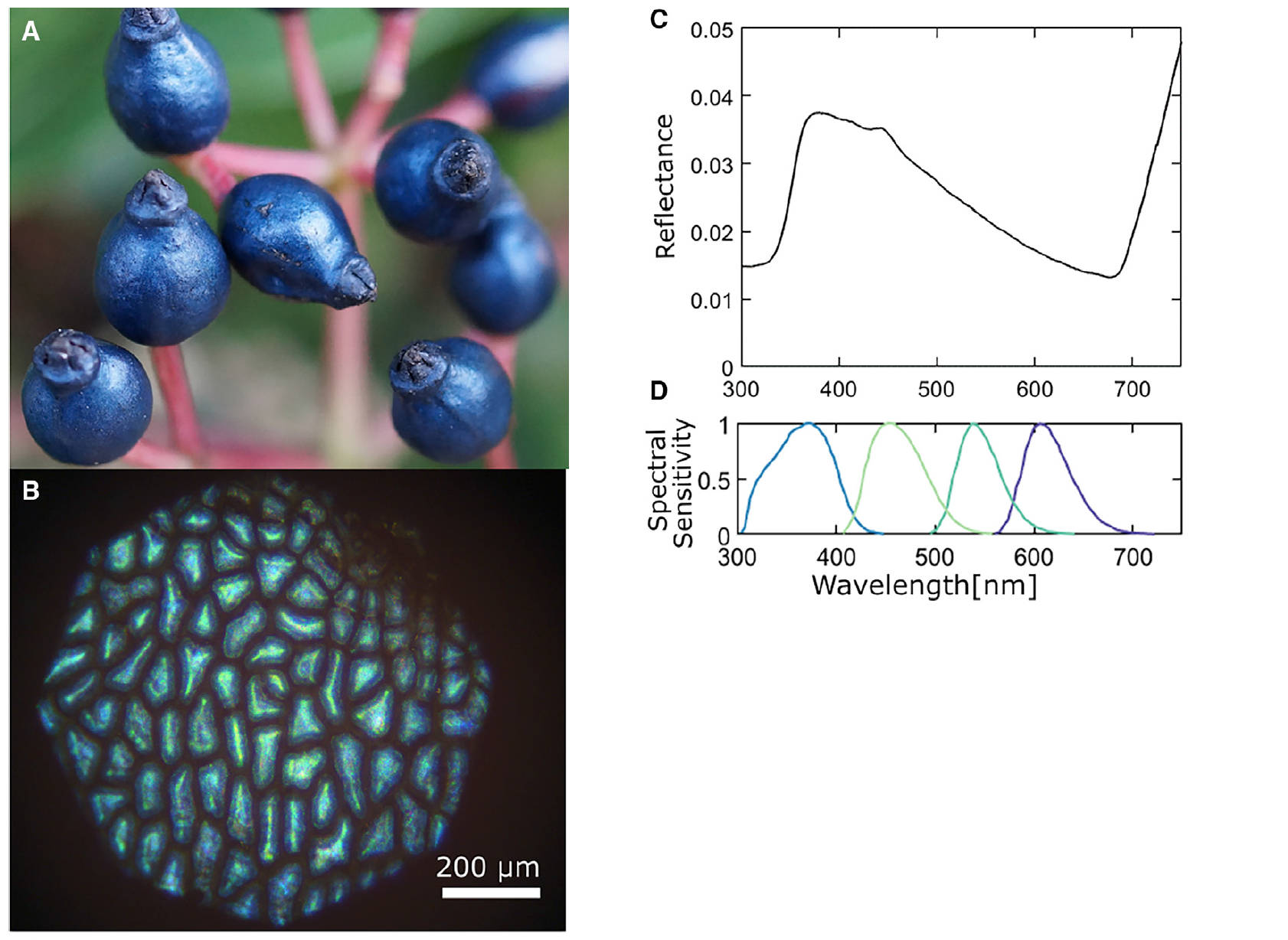
Image n ° 1
Fruits de V. tinus ( 1C) réfléchissent la lumière de manière directionnelle (lui donnant un aspect métallique) dans les régions bleue et ultraviolette du spectre. La polarisation de la lumière réfléchie est largement conservée, indiquant que la coloration est structurelle et non pigmentée, résultant de la réflexion de la paroi cellulaire hautement structurée de l' épicarpe externe * ( 2A ).
L'épicarpe * est la couche externe du fœtus.Lorsque ce tissu est disséqué, un pigment anthocyanique rouge foncé est libéré. La lumière qui n'est pas réfléchie par la structure photonique est absorbée par le pigment rouge foncé en dessous ( 2A et 3C ).
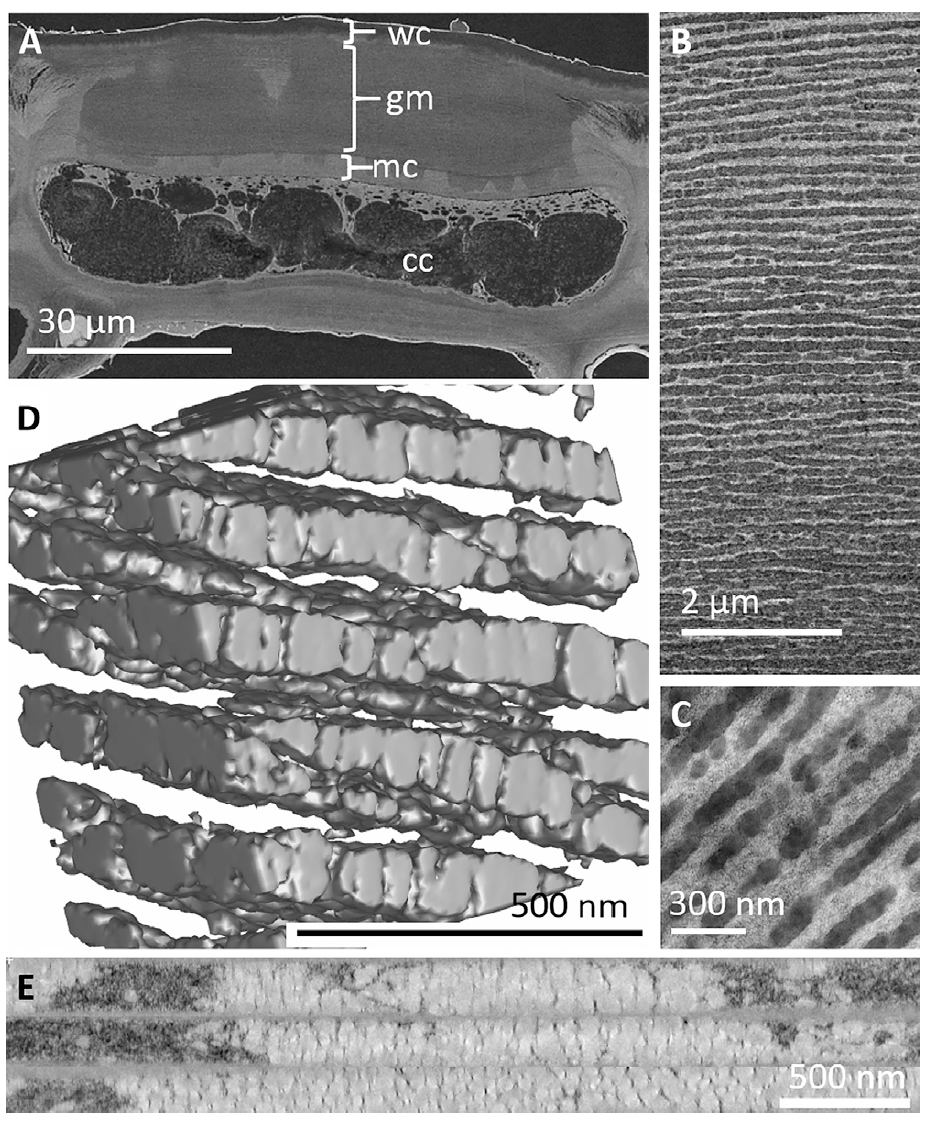
Image # 2
Cette absorption empêche la rétrodiffusion de la lumière, augmentant la visibilité de la réflexion bleue de la paroi extérieure de la cage et améliorant ainsi visuellement la coloration bleue.
À partir de ces observations, on peut déjà conclure que la couleur des fruits de V. tinus est le résultat d'une combinaison d'une nanostructure physique qui réfléchit sélectivement les ondes de lumière bleue et d'une couche de base de pigments améliorant le bleu. En d'autres termes, la conjonction de la chimie et de la physique.
Pour caractériser les nanostructures qui créent la couleur bleue des fruitsV. tinus , les scientifiques ont utilisé plusieurs méthodes de microscopie électronique.
La microscopie électronique à balayage de tissu frais ( 2A ) montre clairement la présence d'une structure multicouche épaisse (10-30 um) parallèle à la surface fœtale et intégrée dans la paroi cellulaire des cellules épicarpiennes les plus externes. La surface du fruit est recouverte d'une cuticule cireuse (2–6 µm) sur une structure en couches. L'architecture en couches occupe la majeure partie de la paroi cellulaire externe dans la zone située entre la cuticule et la paroi cellulaire primaire riche en cellulose. Les couches ont une épaisseur de 30 à 200 nm et recouvrent toute la cellule.
La microscopie électronique à transmission montre que cette architecture est constituée de nombreuses couches de petites bulles, qui diffèrent de la matrice par leur capacité de diffusion des électrons et leur indice de réfraction.

Image # 3 Les
images de microscopie à balayage et à transmission montrent que la matrice semble contenir des composants clés des parois cellulaires végétales typiques, à savoir la cellulose, l'hémicellulose et la pectine. La coloration au rouge ruthénium ( 3D ) montre une teneur significative en pectine, et un diagramme de diffraction électronique démontre la présence de cellulose sur les anneaux de diffraction caractéristiques d'un cristal de cellulose naturelle.
Il est à noter que les couches contrastées sont discrètes et restent distinctes les unes des autres, mais un désordre important est introduit par des couches adjacentes non parallèles et l'inégalité de leur structure globulaire.
L'imagerie de la paroi cellulaire épidermique ( 2E ) montre que ces vésicules globulaires sont organisées en couches combinées à travers lesquelles la matrice de paroi cellulaire de cellulose reste reliée par des ponts et des filaments ( 2B et vidéo ci-dessous).
Modèle de structure multicouche globulaire (correspond à une image 2D ).
Il en découle que la structure multicouche globulaire de l'épiderme des fruits de V. tinus est constituée de lipides noyés dans la matrice de la paroi cellulaire selon différentes méthodes.
Ensuite, des tranches ultrafines de l'épiderme du fruit ont été exposées au chloroforme. Cette analyse est très indicative, car la solubilité dans les solvants organiques non polaires est une indication claire de la présence de lipides.
Images TEM de la même zone d'échantillonnage avant ( 3A ) et après ( 3Ba) l'exposition au chloroforme indique que la structure globulaire a été éliminée par traitement. Dans la dernière image, le contraste de la phase globulaire multicouche est réduit, tandis que les structures vides au sein de la matrice restent visibles. En comparaison, l'exposition à l'eau n'a pas modifié l'ultrastructure ou le contraste d'image du stratifié globulaire, ce qui indique que le matériau ne peut être extrait qu'avec des solvants non polaires. De plus, lorsque les scientifiques ont utilisé du tétroxyde d'osmium tamponné à l'imidazole (C 3 H 4 N 2 / OsO 4 ), qui se lie aux lipides, dans le processus de fixation chimique , les couches globulaires se sont tachées, ce qui confirme leur nature lipidique.
Et lorsque le rouge de ruthénium, qui se lie à la pectine, a été utilisé, la matrice de la paroi cellulaire a été colorée, tandis que la structure globulaire a été éliminée en raison de l'absence de tampon imidazole.
Pendant toutes les variations de coloration appliquées au cours de l'étude, des contours sombres ont été observés autour des globules ( 3E ). Selon les scientifiques, cela peut indiquer la présence d'une membrane lipidique, théoriquement nécessaire à l'interface entre les molécules hydrophobes et les polysaccharides hydrophiles de la paroi cellulaire secondaire.
Les scientifiques nous rappellent que les lipides sont composés d'une variété de structures moléculaires, généralement classées comme cires, graisses et huiles, en fonction de leur point de fusion.
À la surface de l'épiderme des plantes, des cires peuvent facilement être trouvées, formant une cuticule cireuse imperméable. Les cires comprennent également de nombreuses structures moléculaires, mais le composant prédominant est les alcanes, qui sont pratiquement indigestes, c'est-à-dire n'ont aucune valeur nutritive pour les oiseaux. Les graisses et les huiles, en revanche, sont des composants nutritionnels vitaux, car elles contiennent beaucoup plus d'énergie par unité de volume que l'amidon ou les protéines. Les graisses peuvent généralement être trouvées en grandes quantités dans les graines, c.-à-d. profondément à l'intérieur du fœtus.
Dans le cas des fruits de V. tinusLa proximité de la structure globulaire à la fois de grandes graines riches en énergie et de la cuticule externe cireuse rend la distinction entre la cire et les graisses particulièrement importante pour comprendre la signification fonctionnelle et l'origine de la structure. Par conséquent, il est nécessaire de déterminer si les globules lipidiques sont de la cire non digestible ou des graisses et huiles nutritives. Pour cela, la microscopie optique a été utilisée.
Des coupes de tissus de V. tinus foetus ont été incubées avec du bleu du Nil A (pigment), qui colore la région riche en globules de la paroi cellulaire de V. tinus en bleu ou bleu-violet ( 3C). Cela suggère que les globules sont des acides gras libres, et non le polymère de la cutine (un composant de la membrane cuticulaire), qui deviendrait rose ou rouge.
De plus, le diagramme de diffraction électronique de la structure globulaire multicouche montre un motif annulaire clair, qui diffère du diagramme de la paroi cellulaire de cellulose avec les deux anneaux caractéristiques de cristaux de cellulose. Ce modèle indique que les corps lipidiques sont probablement cristallins et sont donc des lipides monomères homogènes plutôt que des molécules polymérisées telles que la cutine.
Pour confirmer que la structure mixte observée de la matrice de cellulose et des globules lipidiques en couches est responsable de la réflectivité bleue des fruits de V. tinus, les scientifiques ont modélisé sa réponse optique. Pour cela, les scientifiques ont étudié deux modèles mathématiques: un tableau bidimensionnel de sphères et une moyenne sur un ensemble de multicouches bidimensionnelles unidimensionnelles.
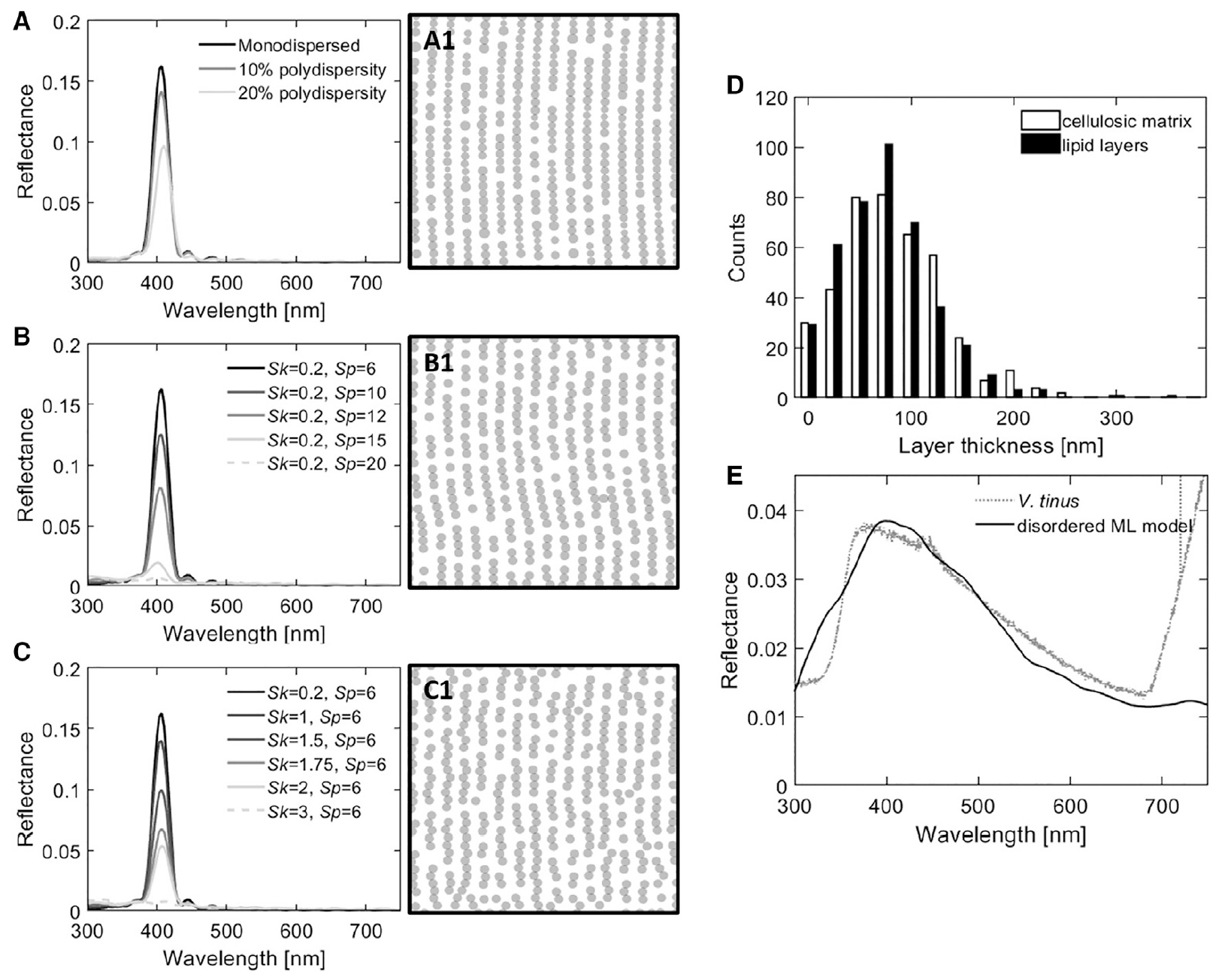
Image # 4
Un algorithme d'ingénierie inverse a été utilisé pour modéliser la structure dans un espace bidimensionnel sous la forme d'une série d'amas globulaires. Les schémas 4A - 4C correspondent au spectre de réflectance modélisé adjacent. Cet algorithme permet à différents types de troubles d'être introduits indépendamment dans la multicouche globulaire en ajustant la taille et la structure, c.-à-d. Transformée de Fourier de la position des particules.
Dans le processus de modélisation, les éléments suivants ont été étudiés: la réponse optique des globules lipidiques en couches avec des degrés variables de variation du diamètre des globules ( 4A); désordre dans les angles entre globules adjacents (paramètre Sp, 4B ) et désordre à la distance moyenne entre globules adjacents (paramètre Sk, 4C ).
L'introduction de différents types de troubles ( 4A - 4C ) a toujours eu le même effet sur la réponse optique de la multicouche globulaire, à savoir, une diminution de l'intensité du pic.
Ainsi, au lieu de considérer chaque élément de trouble séparément, la structure et la composition du matériau de la paroi cellulaire de V. tinus ont été approximées par des multicouches unidimensionnelles désordonnées avec des indices de réfraction correspondant à la cellulose (n = 1,55) et aux lipides végétaux typiques (n = 1,47). La distribution d'épaisseur des deux matériaux est indiquée dans4D . Et la réflectivité, modélisée à l'aide de moyennes sur des couches 1D, est représentée sur la figure 4E .
L'introduction du désordre observé dans les mesures de section transversale dans le modèle de réflecteur ordonné cohérent élargit son spectre de réflexion.
Si le modèle permettait aux scientifiques de comprendre exactement comment les baies de V. tinus obtiennent leur couleur, la modélisation ne couvre pas la nécessité d'un tel mécanisme.
La plus grande interaction interspécifique chez V. tinus est associée aux oiseaux qui se nourrissent des baies de cette plante étonnante. Comparaison avec la sensibilité spectrale de la mésange ( 1D) montre que la couleur des baies se situe dans la fourchette visuellement significative pour les oiseaux de cette espèce.
Les baies, bien sûr, ne flottent pas dans l'air, mais sont attachées aux branches sur lesquelles poussent les feuilles - un arrière-plan visuel. Dans une large mesure, le fond est vert en raison du pigment chlorophylle dominant dans les feuilles. La chlorophylle a une large caractéristique spectrale, avec un pic à 550 nm et une faible réflectivité en dessous de 500 nm, ce qui donne à la couleur des fruits de V. tinus un contraste chromatique avec le feuillage. En d'autres termes, les baies sont encore plus visibles sur le fond de ces feuilles.
Étant donné que les signaux visuels sont souvent prioritaires pour les oiseaux, la coloration structurelle lipidique des baies de V. tinus peut servir de signal visuel fort pour les oiseaux affamés.
Étant donné que la couleur des aliments pour les oiseaux peut être le principal paramètre de comestibilité, la couleur des baies de V. tinus indique qu'elles sont comestibles et nutritives.
La relation entre la couleur du fruit et sa valeur nutritionnelle a été étudiée précédemment. Selon certains rapports, les fruits noirs des plantes de la région brésilienne sont riches en glucides et les fruits noirs des plantes méditerranéennes sont riches en lipides.
Les scientifiques pensent que dans le cas de V. tinus, la couleur bleue est un signal que les baies sont riches en lipides nutritifs, qui, d'ailleurs, créent cette couleur.
Les scientifiques appellent cette méthode de signalisation «honnête» ou «directe» lorsque le contexte du signal correspond à sa source (couleur bleue due aux lipides - teneur élevée en lipides). Cette méthode de signalisation est assez coûteuse, car l'utilisation de la pigmentation classique serait plus facile pour la plante. Néanmoins, le retour que V. tinus obtient en attirant l'attention des oiseaux de différentes espèces, semble-t-il, surmonte cet inconvénient.
Pour une connaissance plus détaillée des nuances de l'étude, je vous recommande de vous pencher sur le rapport des scientifiques .
Épilogue
La couleur est une composante importante des informations visuelles que les organismes vivants reçoivent sur le monde qui les entoure. De nombreux animaux utilisent leur couleur pour se camoufler, attirer des partenaires ou effrayer leurs ennemis. Certaines de ces tactiques sont également présentes dans les plantes, mais la plus importante est le maintien de la communication interspécifique. Dans le cas de V. tinus, les oiseaux sont les principaux partenaires de cette plante, nécessaires à l'épandage des graines sur de longues distances, ce qui augmente considérablement l'habitat de V. tinus et, par conséquent, les chances de survie de l'espèce.
Le goût des fruits de nombreuses plantes dépend de leur volonté d'attirer l'attention des animaux de diverses espèces. Certains fruits seront savoureux, par exemple, pour certaines espèces d'oiseaux, tandis que pour tous les autres, ils seront pratiquement non comestibles. Dans un système aussi complexe que la communication interspécifique, un rôle important est joué par le degré de co-évolution des espèces végétales et animales qui le composent.
La couleur bleue du laurier viburnum réside dans son origine non standard - les nanostructures lipidiques contenues dans les parois des cellules épidermiques des baies de V. tinus . Cette méthode de coloration (structurelle), notamment due aux lipides, n'a jusqu'à présent été trouvée chez les plantes que chez V. tinus... De plus, la coloration lipidique peut indiquer aux oiseaux une teneur élevée en lipides dans les baies, aussi étrange que cela puisse paraître.
Les signaux honnêtes, dont l'origine correspond à leur contexte, sont de nature assez rare. L'explication de cette rareté est assez simple. Imaginez que vous possédez une boulangerie. Vous voulez attirer plus de clients, vous distribuez donc des flyers. Par conséquent, le signal a un contexte (nous avons de délicieux petits pains), mais son origine est différente (je n'ai pas de petit pain, mais quelque chose de mieux est un dessin d'un petit pain, c'est-à-dire que le dépliant n'est qu'un morceau de papier). Si vous distribuez des petits pains, ce sera une signalisation honnête, mais beaucoup plus chère.
Architectures lipidiques précédemment multicouches comme les baies de V. tinusn'ont pas été vus dans le biomatériau. Dans le passé, il n'existait pas d'outils et de techniques aussi avancés que maintenant, par conséquent, de nombreux détails étaient enregistrés de manière incorrecte ou manquaient complètement.
À l'avenir, les scientifiques ont l'intention d'analyser d'autres plantes qui, en théorie, peuvent également avoir des nanostructures lipidiques similaires et, par conséquent, une méthode non standard pour la coloration des fruits. De plus, les scientifiques pensent que leurs recherches pourraient aider à créer des colorants alimentaires plus sûrs.
Merci pour votre attention, restez curieux et passez un excellent week-end, les gars! :)
Un peu de publicité
Merci de rester avec nous. Aimez-vous nos articles? Vous voulez voir du contenu plus intéressant? Soutenez-nous en passant une commande ou en recommandant à des amis, le cloud VPS pour les développeurs à partir de 4,99 $ , un analogue unique des serveurs d'entrée de gamme que nous avons inventé pour vous: Toute la vérité sur VPS (KVM) E5-2697 v3 (6 cœurs) 10 Go DDR4 480 Go SSD 1 Gbps à partir de 19 $ ou comment diviser correctement le serveur? (options disponibles avec RAID1 et RAID10, jusqu'à 24 cœurs et jusqu'à 40 Go de DDR4).
Le Dell R730xd 2x est-il moins cher dans le centre de données Equinix Tier IV à Amsterdam? Seulement, nous avons 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2,6 GHz 14C 64 Go DDR4 4x960 Go SSD 1 Gbps 100 TV à partir de 199 $ aux Pays-Bas!Dell R420 - 2 x E5-2430 2,2 GHz 6C 128 Go DDR3 2 x 960 Go SSD 1 Gbps 100 To - À partir de 99 $! Lisez à propos de Comment construire l'infrastructure de bldg. classe avec les serveurs Dell R730xd E5-2650 v4 au coût de 9000 euros pour un centime?