
En tant que personne aux dimensions non standard (à la vadrouille avec oreilles), je sais parfaitement combien il est parfois difficile de choisir n'importe quel article de la garde-robe de la bonne taille. Apparemment, certains des scientifiques de l'Université de Harvard ont également été confrontés à un problème quotidien similaire, car dans leurs recherches récentes, ils décrivent un nouveau type de matériau à mémoire de forme. La base de cette innovation est une protéine que l'on trouve facilement dans les cheveux, les ongles et la peau - la kératine. Comment exactement le favori des spécialistes du marketing des fabricants de cosmétiques a-t-il été utilisé, de quel type de métamorphose le nouveau matériau est-il capable et quelles sont les options pour utiliser le matériau métamorphique? Pour obtenir des réponses à ces sondages, plongeons-nous dans le rapport des scientifiques. Aller.
Base de recherche
Au cours des dernières années, l'intérêt pour les matériaux capables de se souvenir de certaines formes données lors de la production s'est considérablement accru. Ces matériaux peuvent être utilisés dans les industries de la médecine, de la construction et de l'aérospatiale, etc. Cependant, comme le déclarent les scientifiques eux-mêmes, le degré d'intérêt pour de tels développements ne peut être assimilé au degré d'informations disponibles à leur sujet. En d'autres termes, ces matériaux ont encore de nombreux secrets non résolus.
Les matériaux métamorphiques sont le plus souvent associés à des substances synthétiques; cependant, les structures naturelles ont également des propriétés similaires, ce qui est dû à la métastabilité structurelle des structures secondaires des protéines. Par exemple, les hélices α de kératine, disposées en bobine hélicoïdale, sont connues pour subir une transition structurelle continue en feuillets β métastables lorsqu'une charge est appliquée le long de leur axe longitudinal. Selon le type d'α-kératine, ce processus peut être irréversible ou réversible, et dans le second cas il ressemble au mécanisme martensitique * à mémoire de forme des alliages métalliques.
La transformation martensitique * est une transformation polymorphe dans laquelle un changement dans l'arrangement mutuel des atomes (ou molécules) d'un cristal se produit par leur mouvement ordonné.Dans les matériaux biologiques (peau animale par exemple), un tel mécanisme de transformation est dû à la nécessité d'une protection et d'un fonctionnement physiologique en réponse à un stimulus externe.
Dans ce travail, les scientifiques ont tenté de mettre en œuvre la transition réversible de la kératine de l'hélice α à la feuille β. Selon eux, c'est ce processus qui est le mécanisme principal pour créer un matériau nanostructuré de haute technologie à mémoire de forme, qui utilise l'hydratation comme déclencheur et est de nature biocompatible et biodégradable.
Résultats de recherche
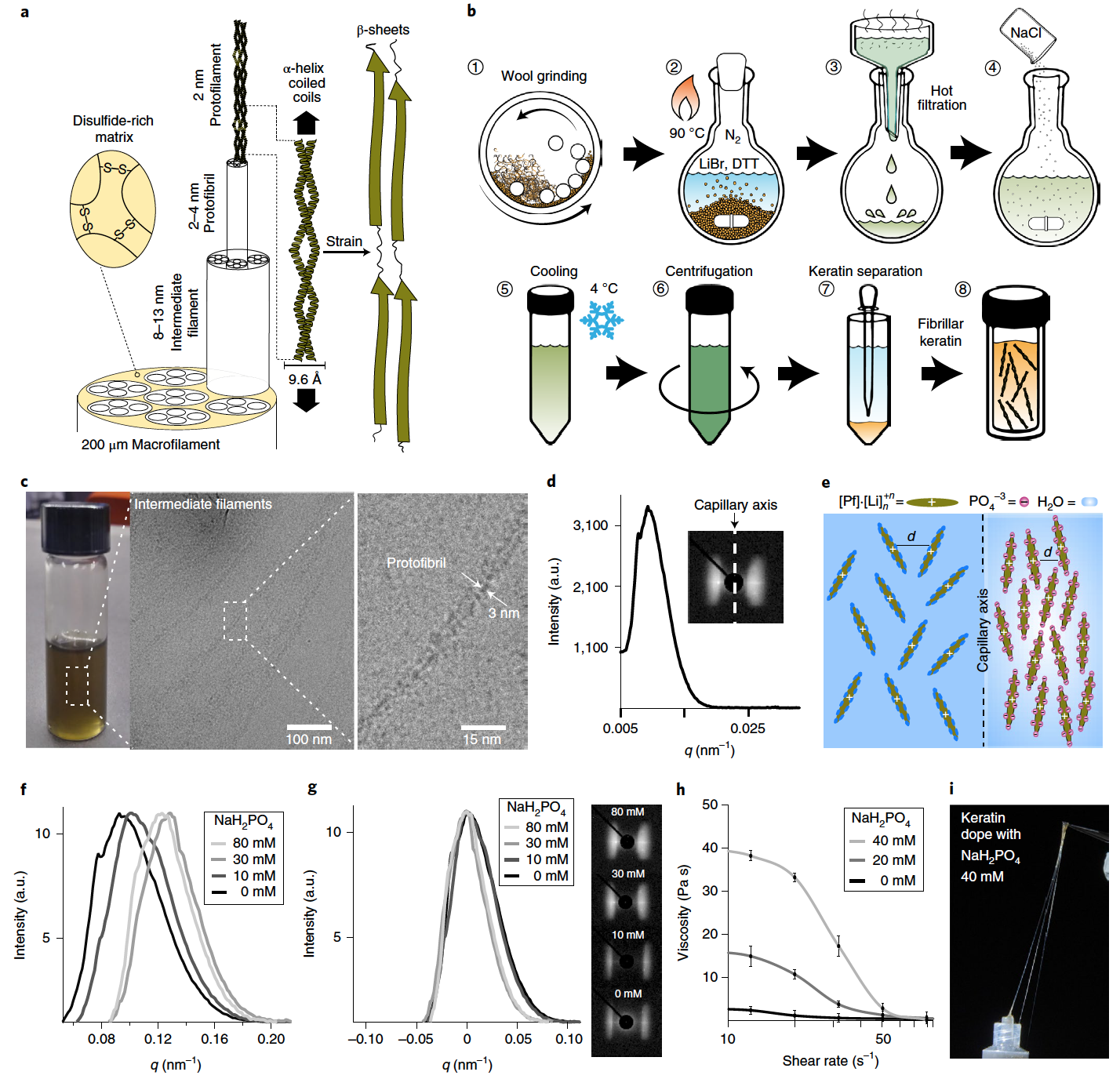
Image n ° 1
Dans les poils d'animaux, la transition induite par la déformation de l'hélice α à la feuille β est possible grâce à la configuration appariée des hélices α dans l'architecture de la bobine en spirale * ( 1a ).
Spiral coil * ( Coiled coil ) - protéines de motifs structuraux, lorsque 2 à 7 hélices alpha entrelacent les deux brins de la corde.Les bobines en spirale s'auto-organisent hiérarchiquement en une structure fibrillaire anisotrope qui va des protofibrilles * aux macrofibrilles * , ce qui assure la continuité de la transformation mécanique à toutes les échelles spatiales.
Les protofibrilles * sont les filaments de protéines les plus minces qui constituent l'essentiel des myofibrilles.Dans cette étude, la kératine fibrillaire a été extraite de la laine d'angora à l'aide de bromure de lithium (LiBr), un sel qui peut induire une transition réversible de phase solide à liquide de la kératine cristalline dans l'eau.
Myofibrilles * - filaments contractiles dans le protoplasme des fibres musculaires striées des muscles squelettiques, du muscle cardiaque et des muscles à double striation oblique.
Une autre exigence pour la libération de kératine fibreuse de la structure capillaire est la rupture du réseau disulfure dense du composant de matrice capillaire. Ceci a été réalisé en utilisant 1,4-dithiothréitol (DTT, C 4 H 10 O 2 S 2 ), qui est capable de cliver la liaison disulfure pour former deux fractions sulfhydryle. Cette réaction est réversible dans des conditions d'oxydation, ce qui permet de restaurer les ponts disulfure naturels lors de la fabrication.
La kératine a ensuite été récupérée avec succès en traitant la laine avec une solution aqueuse de LiBr et de DTT à haute température ( lb ). A température ambiante, la kératine a finalement été isolée par séparation liquide, conduisant à une solution de kératine hautement concentrée avec une durée de conservation de plusieurs semaines en l'absence d'oxygène.
La spectroscopie Raman et le dichroïsme circulaire ont également été réalisés, ce qui a confirmé la présence de bobines hélicoïdales d'hélices α. La preuve d'une structure hiérarchique de la kératine jusqu'au niveau protofibrillaire a été confirmée par microscopie électronique à transmission cryogénique (cryo-TEM). Au cours de la microscopie, il a été constaté que la longueur des nœuds est à quelques micromètres et la largeur est d'environ 10 nm ( 1c), qui correspond pleinement aux caractéristiques structurelles des fibres intermédiaires. Il a également été possible d'établir que la structure hiérarchique des filaments intermédiaires est constituée de protofibrilles tassées d'une largeur constante de ~ 3 nm.
La mise en œuvre d'une architecture de kératine hiérarchique qui peut fournir une commande à longue distance des unités exécutives à hélice α nécessite l'imposition d'un alignement anisotrope des protofibrilles pendant le processus de fabrication. Il a été constaté que les protofibrilles de kératine s'auto-assemblent en une phase cristalline nématique sous l'influence de la contrainte de cisaillement et des contraintes spatiales. Ce fait a été établi en observant la nature anisotrope de la diffusion des rayons X synchrotron, qui a été obtenue à partir d'un échantillon de solution de kératine (401,7 mg / ml) préparé dans un capillaire de quartz ( 1d).
Au cours de l'analyse, le capillaire était positionné perpendiculairement au faisceau de rayons X et son axe longitudinal était parallèle à l'axe méridien du détecteur. Le caractère équatorial de la diffusion suggère que les domaines de la kératine étaient principalement orientés parallèlement à l'axe capillaire (encart en 1d ). La distance moyenne entre les domaines de la kératine est liée au paramètre de taille de réseau ( d ), qui est obtenu à partir de l'intensité maximale du module de vecteur de diffusion ( q ): d = 2 π / q .
Les scientifiques suggèrent que l'ordre nématique du protofilament de kératine * est le résultat de la contrainte de cisaillement qui est créée sur la paroi capillaire pendant la préparation de l'échantillon et est également le résultat d'une stabilisation supplémentaire en raison de l'espace limité.
Les protofilaments * sont des structures protéiques filamenteuses qui sont des éléments constitutifs des microtubules (structures intracellulaires protéiques qui composent le cadre cellulaire).Dans un tel cas, on s'attend à ce que la rigidité accrue et l'auto-assemblage des protofibrilles de kératine se traduisent par un degré d'ordre plus élevé dans la phase nématique (filamenteuse). Le contrôle de l'auto-organisation de la phase de cristaux liquides de la kératine a été obtenu en stimulant les interactions protéine-protéine par un effet de filtrage de charge. En raison de la présence de cations lithium, qui sont absorbés à la surface de la protéine, la kératine aura une charge positive nette. Et l'anion phosphate a été utilisé car il a un effet d'écran élevé vis-à-vis des surfaces chargées positivement ( 1e ).
L'ajout de phosphate monosodique (NaH 2 PO 4) a provoqué un resserrement du tassement de la phase nématique de la kératine, ce qui est indiqué par le déplacement du pic vers une valeur q ( 1f ) plus élevée .
L'ajout de sel kosmotrope * a également entraîné un rétrécissement du motif de diffusion équatoriale et, par conséquent, une exacerbation du pic de diffusion, ce qui indique une augmentation de l'alignement du domaine de la kératine le long de l'axe capillaire ( 1g ).
Les cosmotropes * sont des co-solvants s'ils contribuent à la stabilité et à la structure des interactions eau-eau. Les cosmotropes provoquent une augmentation de l'interaction des molécules d'eau, ce qui stabilise également les interactions intramoléculaires dans les macromolécules telles que les protéines.Avec une augmentation de la concentration de NaH 2 PO 4, l' agrégation des protofibrilles provoque une augmentation de la viscosité de la solution protéique à de faibles taux de cisaillement (1 h ). Cependant, avec l'augmentation du taux de cisaillement, l'alignement des protofibrilles de kératine provoque une diminution soudaine de la viscosité, ce qui confère à la solution protéique une pseudoplasticité prononcée * .
Pseudoplasticité * - se produit lorsque la viscosité d'un fluide diminue avec l'augmentation de la contrainte de cisaillement.A une concentration de NaH 2 PO 4 de 40 mM et une concentration de protofilament de 401,7 mg / ml, le dopant kératine présente des propriétés viscoélastiques. C'est une bonne nouvelle pour les chercheurs, car les fibres peuvent être formées directement en tirant simplement sur la protéine avec une pince à épiler ( 1i ). Si la concentration de NaH 2 PO 4 est réduite , la solution de kératine perd ses propriétés viscoélastiques et ne peut pas former de fibres directement à partir de la solution.
Comme l'ont noté les scientifiques, l'alignement des hélices α de kératine le long de l'axe de la fibre est un critère de conception qui garantit une résistance élevée et un degré élevé de fixation de la fibre. Lorsque les axes de l'hélice α sont parallèles au vecteur de tension, un dénouement maximal des hélices α peut être obtenu, ce qui permet au matériau d'augmenter la déformation avant rupture due à la déformation plastique et à la réorganisation.

Image n ° 2
Une solution aqueuse de NaH 2 PO 4 a été utilisée comme anti-solvant, ce qui a permis d'obtenir une diffusion externe de LiBr à partir du dopant à la kératine extrudé et un auto-assemblage supplémentaire de la protéine en raison de l'effet de filtrage de charge ( 2a). La réduction du réseau covalent disulfure est devenue possible grâce à l'activité oxydante du peroxyde d'hydrogène (H 2 O 2 ) vis-à-vis du groupe thiol de la cystéine. La concentration élevée de protéines dans le dopant confère à la fibre la résistance pendant le processus de coagulation, ce qui permet un processus de filage flexible et fiable.
Il en résulte que des fibres longues et résistantes ( 2b ) peuvent être obtenues , et la vitesse de production élevée permet d'obtenir des fibres d'un diamètre de 10 µm.
L'organisation de la phase nématique des protofibrilles de kératine entraîne un processus de fibrillation qui génère des fibres structurées hiérarchiquement et anisotropes. Le microscope électronique à balayage a montré qu'une fibre est constituée de fibrilles continues dont la longueur est d'au moins plusieurs dizaines de micromètres ( 2b ). Il a également été observé que les fibrilles d'un diamètre de 50 nm sont le cœur de la fibre résultante ( 2d ).
La microscopie optique polarisante a confirmé la nature anisotrope du cœur de la fibre, qui a été établie en observant la biréfringence avec une intensité lumineuse maximale transmise à un angle de 45 ° ( 2e ).
La structure de la bobine en spirale a une architecture anisotrope, elle a été établie en utilisant la diffusion des rayons X grand angle (WAXS ). Le profil de diffusion bidimensionnel montre une réflexion équatoriale caractéristique à 9,65 Â, qui correspond à la distance entre les axes des hélices α adjacentes ( 2g ). L'analyse unidimensionnelle le long de l'axe méridien montre la présence de réflexions méridiennes caractéristiques (à 5,15 Å) et extra-méridiennes (5,05 Å), qui correspondent à la projection du pas de l'hélice α ( 2h ).
Il a également été constaté que le maximum est décalé vers des valeurs q plus élevées , c.-à-d. il y a des chaînes peptidiques dépliées orientées parallèlement à l'axe de la fibre et formant probablement une conformation en couche β (2i ).
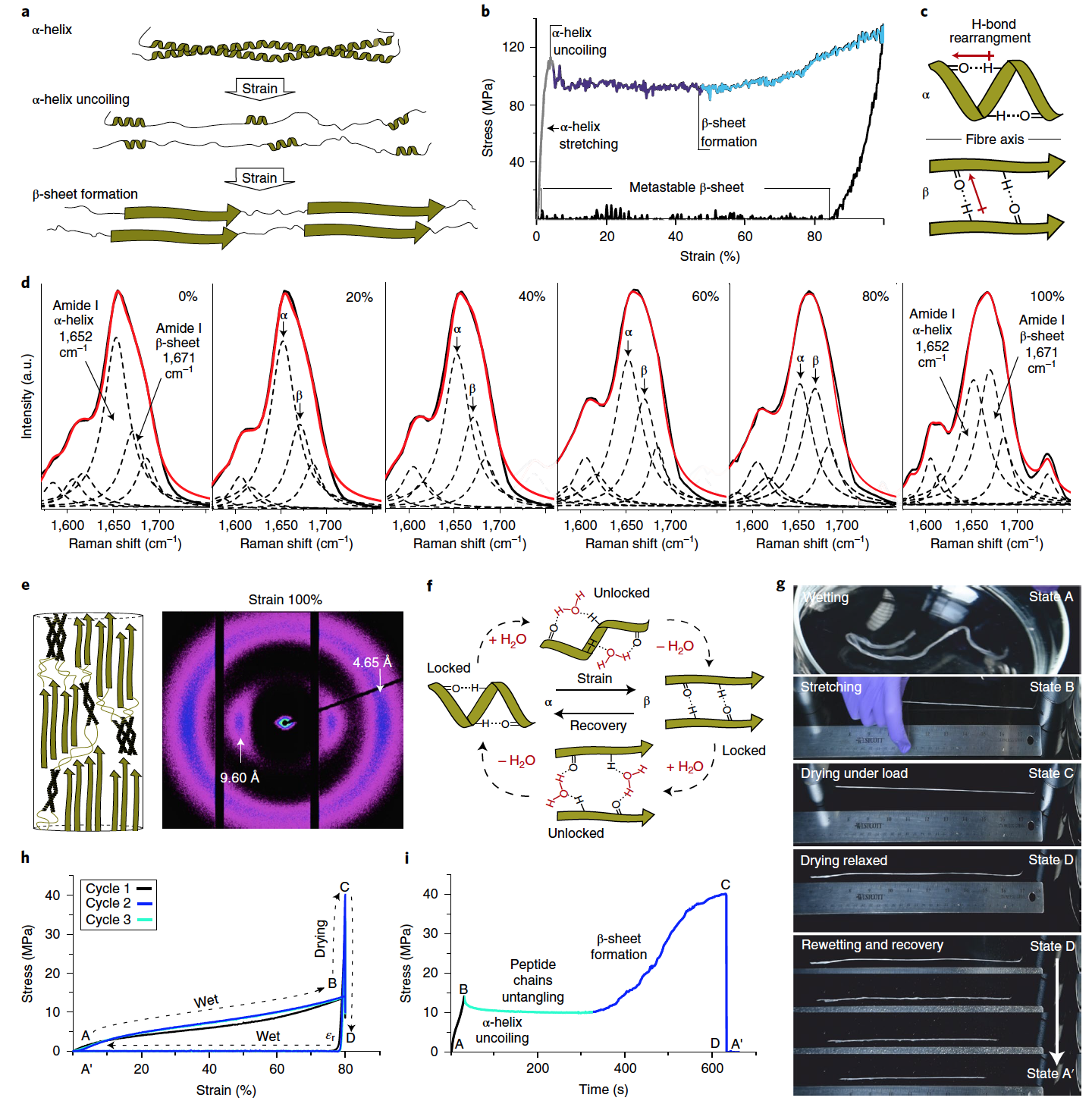
Image №3
Lors de la prochaine étape de l'étude, nous avons étudié les fibres à mémoire de forme qui répondent à l'hydratation.
L'effet mémoire de forme des fibres kératiniques artificielles est basé sur le déroulement réversible de l'hélice α et la formation de feuillets β métastables en déformation uniaxiale ( 3a ).
Des tests de traction réalisés sur des fibres kératiniques individuelles ont confirmé ce mécanisme. L'état élastique initial a été établi jusqu'à ~ 5% de déformation (module de Young = 4,18 ± 0,10 GPa). Ceci est suivi d'une région caractérisée par une limite élastique constante (96,1 ± 3,1 MPa) ( 3b ). Cette réaction correspond au processus de dépliage de l'hélice α.
Au fur et à mesure que le stress augmente, les chaînes peptidiques dépliées et étendues de la kératine sont stabilisées dans leur géométrie étendue par assemblage en couches β ( 3c ). Cette région de formation de la feuille β est caractérisée par un durcissement sous contrainte à une déformation d'environ 50%, car la charge appliquée n'est pas seulement dissipée en raison de la destruction des bobines enroulées, mais est également transférée par étirement des feuilles β. Lorsque la charge est supprimée à une déformation de 100% (résistance à la traction 137,18 ± 1,03 MPa), la fibre présente une déformation plastique (~ 85%), ce qui est cohérent avec la transformation des chaînes kératiniques dépliées en nouvelles feuilles β métastables. Les propriétés mécaniques des fibres kératiniques obtenues correspondent parfaitement à celles de la laine naturelle.
Il est important de noter que les fibres non sollicitées contiennent également une proportion insignifiante de feuillets β ( 3d ); cependant, à 100% de déformation, on observe une augmentation significative du composant en feuillets β. L'analyse WAXS a confirmé la présence d'une transition induite par la déformation de l'hélice α à la couche β ( 3e ).
Dans les fibres étirées, les feuilles β sont cinétiquement stables en raison de la présence d'un réseau de liaisons hydrogène, ce qui les empêche de se reconvertir en hélices α plus stables thermodynamiquement. C'est cette propriété qui permet de créer un système à cycle à mémoire de forme, dans lequel un réseau de liaisons hydrogène joue le rôle d'un mécanisme de blocage pour assurer la fixation d'une forme déformée.
Au cours des expériences, l'eau a joué le rôle d'un stimulus contribuant à la déformation de la fibre et à la restauration de sa forme d'origine ( 3f ). La technique a été testée sur un faisceau de fibres kératiniques de même diamètre ( 3g et vidéo ci-dessous).
Démonstration de la mémoire de forme à l'aide de l'exemple de fibres individuelles.
Tout d'abord, le faisceau de fibres a été hydraté dans de l'eau désionisée pendant quelques secondes (état A), puis étiré manuellement à l'air alors qu'il était encore humide (état B), puis maintenu sous charge à température ambiante pendant 10 minutes pour permettre aux fibres de sécher ( état C).
Après avoir retiré les poids, ce qui a permis aux fibres d'entrer dans un état détendu, il n'y avait pas de changement visible ou perceptible de longueur entre les formes étirées et relâchées (état D).
Si, cependant, de l'eau est à nouveau appliquée sur les fibres résultantes (par pulvérisation), alors les fibres sont réduites à leur longueur d'origine en quelques secondes (état A ').
L'utilisation d'eau dans les métamorphoses en cours facilite grandement le processus de restructuration de la structure protéique. Ceci est indiqué par une diminution générale de la contrainte de traction ( 3h ) et une transition plus progressive entre les états des fibres.
Lorsque la fibre sèche sous contrainte, la formation d'une feuille β est indiquée par une forte augmentation de la contrainte correspondant à une augmentation de la rigidité de la fibre, qui peut être mesurée avec le temps lorsque les fibres se déshydratent et se lient à l'hydrogène ( 3i ).
Le matériau résultant, en raison de l'ordre à longue portée de sa structure fibrillaire à l'état sec, démontre une résistance à la traction (137,18 ± 1,03 MPa) et un module d'Young (4,18 ± 0,10 GPa), qui est bien meilleur que celui des prototypes précédemment développés. Une fois hydraté, la résistance à la traction est de 14,94 ± 0,46 MPa, ce qui est également nettement supérieur à d'autres matériaux développés.
En plus des propriétés et des caractéristiques, le matériau développé présente un autre avantage sur ses concurrents: la possibilité de son utilisation en impression 3D.

Image n ° 4
Des formes géométriques de base peuvent être obtenues par extrusion du dopant protéique dans un hydrogel, qui sert de bain de support et de coagulation ( 4a). Les propriétés de la kératine permettent l'utilisation de petites aiguilles, ce qui permet la création de structures à une échelle d'environ 50 μm ( 4b ).
L'alignement des protofibrilles de kératine suit le chemin d'extrusion en impression 3D et conduit donc à des architectures très ordonnées qui se caractérisent par une hiérarchie structurelle interne du niveau moléculaire au niveau macroscopique ( 4c ).
Une fois que l'échantillon souhaité a été imprimé en 3D, il est nécessaire d'obtenir une fixation de forme constante. Cela nécessite la formation de ponts disulfure en raison de l'oxydation provoquée par H 2 O 2 . Avant le processus d'oxydation, les échantillons peuvent encore être manipulés en changeant leur forme en raison de leur plasticité.
Par exemple, lors des tests, une étoile (origami) a été fabriquée manuellement à partir de la feuille imprimée, qui a ensuite passé l'étape de fixation de la forme par oxydation dans H 2 O 2 et NaH 2 PO 4 ( 4d ). Par conséquent, il n'est pas nécessaire d'imprimer la forme désirée immédiatement, cela peut être fait après l'impression et avant l'étape de fixation ( 4e ).
Démonstration de la mémoire de formulaires à l'aide de l'exemple d'un échantillon imprimé.
Tout comme les fibres testées précédemment, les figurines imprimées ont les mêmes propriétés de mémoire de forme sensible à l'humidité. L'architecture de l'origami en forme d'étoile a été choisie pour démontrer l'efficacité du mécanisme de mémoire de forme lors de la réalisation de transformations géométriques assez complexes, selon les scientifiques.
Une fois hydraté, le modèle d'origami imprimé en 3D est malléable et peut être déballé et transformé arbitrairement, par exemple, en un tube roulé (à gauche à 4f). En séchant, la feuille carrée perd sa plasticité et se fixe dans sa nouvelle forme provisoire. La restauration de l'architecture de l'origami en forme d'étoile est alors déclenchée par réhydratation, qui se produit en quelques secondes en raison du rapport surface / volume élevé qui fait que la kératine agit rapidement sur l'eau (juste à 4f ). En d'autres termes, la feuille imprimée se déplie d'abord dans sa configuration précédente puis se plie en forme d'étoile.
Pour une connaissance plus détaillée des nuances de l'étude, je vous recommande de consulter le rapport des scientifiques et des éléments supplémentaires .
Épilogue
Dans ce travail, les scientifiques ont mis en évidence un nouveau type de matériau à mémoire de forme, activé par contact avec l'eau. La base du processus de transformation spontanée d'une forme donnée en une autre est la transition des hélices α de la kératine en feuillets β.
Le matériau obtenu peut être utilisé dans l'impression 3D, tandis que vous pouvez initialement définir la forme souhaitée ou le faire après l'impression d'une feuille ordinaire. Le changement de forme après impression est possible en raison de la plasticité de l'échantillon obtenu, dont la forme peut être fixée déjà au stade de l'oxydation. Ce processus en deux étapes permet la création de formes très complexes avec des caractéristiques structurelles personnalisables jusqu'au micron.
Les auteurs de cette étude disent que la gamme d'applications pour leur développement est assez large. Les matériaux à mémoire de forme peuvent être utilisés à la fois dans l'industrie légère (par exemple, un T-shirt qui change de taille à votre guise) et en médecine (tissus activateurs).
Les résultats des expériences sont assez bons, mais les scientifiques ont l'intention de continuer à mener des expériences, car les matériaux, dont l'architecture peut changer en fonction d'influences extérieures et à la demande d'une personne, sont un sujet d'étude très intéressant.
Merci pour votre attention, restez curieux et passez une bonne semaine de travail, les gars. :)
Un peu de publicité
Merci de rester avec nous. Aimez-vous nos articles? Vous voulez voir du contenu plus intéressant? Soutenez-nous en passant une commande ou en recommandant à des amis, Cloud VPS pour les développeurs à partir de 4,99 $ , un analogue unique des serveurs d'entrée de gamme que nous avons inventé pour vous: The Whole Truth About VPS (KVM) E5-2697 v3 (6 Cores) 10GB DDR4 480GB SSD 1Gbps à partir de 19 $ ou comment diviser correctement le serveur? (options disponibles avec RAID1 et RAID10, jusqu'à 24 cœurs et jusqu'à 40 Go de DDR4).
Le Dell R730xd 2x est-il moins cher dans le centre de données Equinix Tier IV à Amsterdam? Seulement, nous avons 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2,6 GHz 14C 64 Go DDR4 4x960 Go SSD 1 Gbps 100 TV à partir de 199 $ aux Pays-Bas!Dell R420 - 2 x E5-2430 2,2 GHz 6C 128 Go DDR3 2 x 960 Go SSD 1 Gbps 100 To - À partir de 99 $! Lisez à propos de Comment construire l'infrastructure de bldg. classe avec l'utilisation de serveurs Dell R730xd E5-2650 v4 au coût de 9000 euros pour un centime?