Le corps humain peut, sans exagération, être classé parmi les systèmes biologiques les plus complexes de la planète. Notre corps se compose de milliards de cellules, de nombreux organes et systèmes. Mais toute cette splendeur et cette diversité sont dues à la fusion de seulement deux cellules - un sperme et un ovule. Je pense qu'il n'est pas nécessaire d'expliquer comment se déroule la fécondation et ce qui est nécessaire pour cela (indice - les cigognes et le chou n'ont rien à voir avec cela). Mais voici quelques aspects de la vie des spermatozoïdes restés flous pendant de nombreuses années. Des scientifiques de l'Université de Bristol, utilisant des techniques modernes de microscopie tridimensionnelle, ont pu voir les mouvements du sperme d'une manière qui n'était pas possible auparavant. Comment et par quels moyens le sperme se déplace, nous apprend le rapport des scientifiques. Aller.
Base de recherche
Bien que les spermatozoïdes aient été impliqués dans la création de la vie bien avant l'avènement de la méthode scientifique, leur chemin dans la littérature scientifique a commencé assez récemment - en 1677. L'étudiant en médecine Johann Gam a partagé ses observations avec son collègue et ami Anthony van Leeuwenhoek (1632 - 1723), qui, à son tour, a examiné et décrit en détail les «animaux séminaux» (comme il les appelait les spermatozoïdes).
Anthony van Leeuwenhoek / Lazzaro Spallanzani / Carl Ernst von Baer
Leeuwenhoek a suggéré que ces cellules inhabituelles sont impliquées dans la fécondation, mais sa théorie, bien que vraie, a été rejetée par la communauté scientifique. Pendant longtemps, on a cru que les spermatozoïdes étaient des parasites et que seul le liquide séminal était impliqué dans la fécondation.
Ce n'est que près de cent ans plus tard, en 1786, que le fait de la participation du sperme à la fécondation fut prouvé par Lazzaro Spallanzani (1729 - 1799) dans son ouvrage "Experiencias Para Servir a La Historia de La Generación De Animales y Plantas". Cependant, ses explications sur le processus lui-même étaient plutôt vagues: il pensait que l'ovule était déjà le début d'un nouvel organisme et que le sperme n'était nécessaire que pour activer le processus de croissance.
Le même terme «sperme» a été introduit au début du 19e siècle par Karl Ernst von Baer (1792 - 1876).
Quoi que pensaient les scientifiques il y a plusieurs siècles, le sperme a une fonction très claire, dont la mise en œuvre est assurée par un certain nombre d'outils spécialisés. La tâche principale du sperme humain est de traverser le tractus génital féminin, de trouver l'ovule et de lui transférer le matériel génétique masculin.

La structure du sperme La
cellule reproductrice masculine ne peut pas se vanter de dimensions, car elle est la plus petite du corps humain (à l'exclusion de la queue): les dimensions de la tête sont de 5,0x3,5x2,5 microns (longueur x largeur x hauteur), la longueur de la partie médiane est de 4,5 microns, et longueur de la queue - 45 microns.
Dans le même temps, la petite taille n'est pas un inconvénient, mais un aspect réfléchi de l'augmentation de sa vitesse. Dans le processus de maturation du sperme, son noyau (porte un seul ensemble de chromosomes) devient plus dense, la majeure partie du cytoplasme est rejetée et seuls les organites les plus importants restent dans la cellule.
Le flagelle peut être appelé le deuxième élément le plus important du sperme après le noyau, c.-à-d. Sa queue. Car c'est grâce à lui que s'effectue le mouvement de cette cellule le long du tractus génital d'une femme. Il est également amusant que l'environnement vaginal soit extrêmement destructeur pour les cellules germinales mâles, mais le sperme réduit partiellement l'effet négatif sur le sperme. Le niveau de pH à l'intérieur du tractus génital féminin permet aux spermatozoïdes de se déplacer vers l'utérus, où un environnement beaucoup plus favorable les attend.
Auparavant, on pensait que le sperme se déplaçait vers l'avant en raison du mouvement symétrique de son flagelle d'un côté à l'autre.
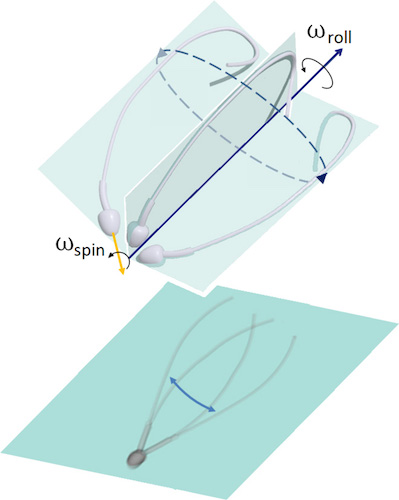
Image n ° 1: asymétrie du mouvement flagellaire en 3D (en haut); projection plane du mouvement du flagelle, créant une illusion d'optique de symétrie bilatérale en microscopie 2D (par le bas).
Cette déclaration a été exprimée même à l'époque de Levenguk. Cela a également conduit à une idéalisation symétrique de la forme d'onde en trois dimensions, souvent perçue comme une spirale conique, semblable à un tire-bouchon en expansion.
En raison de la microscopie bidimensionnelle, de nombreuses observations ont été interprétées de manière inexacte, et parfois complètement incorrecte. Les déclarations concernant la symétrie des lambeaux de flagelle contredisent de nombreuses observations montrant l'asymétrie structurelle dans le cadre du flagelle lui-même.
Si le battement du flagelle du sperme pendant le mouvement est toujours asymétrique, comment la symétrie du mouvement du flagelle d'un côté à l'autre et le mouvement de la cellule vers l'avant sont-ils obtenus? C'est cette question qui est la principale de cette étude.
Pour obtenir une réponse à cela, les scientifiques ont comparé des observations moléculaires et microscopiques qui ont montré que le sperme humain utilise à la fois un contrôle asymétrique et anisotrope pour réguler le battement flagellaire. En d'autres termes, la symétrie est réalisée en raison de l'asymétrie: l'effet de «pré-précession» se produit lorsque la tête tourne («fluide de forage», selon les auteurs) simultanément et indépendamment de la façon dont le flagelle du sperme tourne autour de l'axe de mouvement.
Résultats de recherche
Le mouvement rapide des flagelles de spermatozoïdes humains a été enregistré avec une haute résolution spatiale et temporelle en 3D. Deux groupes de spermatozoïdes flottant librement dans un fluide à faible viscosité ont été examinés: les spermatozoïdes qui flottent à côté de la lamelle (placé sur le dessus de l'échantillon pendant la microscopie) et à l'écart de celui-ci.
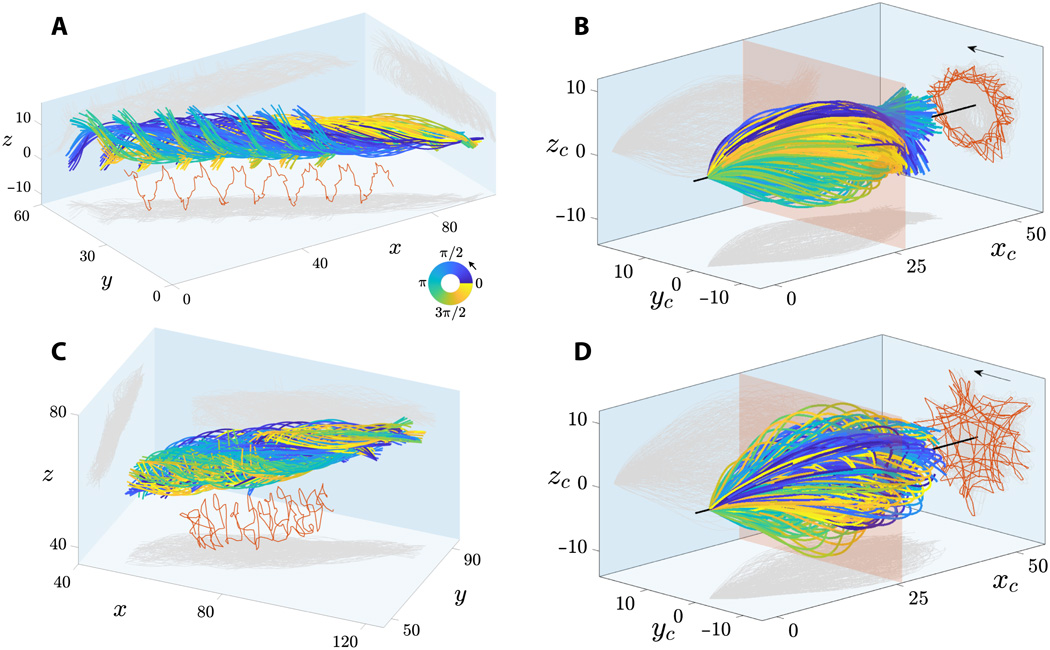
Image №2
Le spectacle 2A et 2C balaie les flagelles de sperme flottant près et loin de la lamelle. Les lambeaux de flagelle sont caractérisés par un mouvement de roulement caractéristique autour de la direction du mouvement du sperme.
La rotation et le mouvement de translation combinés du flagelle du sperme conduisent à des trajectoires en spirale du point médian du flagelle avec une chiralité préservée (marquée en rouge à 2Aet 2C ). Les spermatozoïdes montrent une rotation bidirectionnelle autour de leur axe: toutes les cellules flottant librement (28 pièces) ont tourné dans le sens antihoraire lorsqu'elles sont vues de l'avant (flèches sur 2B et 2D ) et seulement 2 cellules ont tourné dans le sens des aiguilles d'une montre (elles n'ont pas avancé en raison d'obstacles En route).
Microscopie des spermatozoïdes flottant à côté de la lamelle (correspond à 2A ).
Tous les spermatozoïdes (30 cellules) se sont déplacés de la même manière vers un sommet rotatif précédent, dans lequel la rotation de la tête autour de l'axe longitudinal du spermatozoïde (ω spin ) se produit simultanément et indépendamment de la façon dont le flagelle tourne par rapport à l'axe de mouvement (ω roll ).
En 2B et 2D montre des termes de flagelle battant du système de coordonnées concomitant * , c'est-à-dire un point de vue, se déplace avec le sperme, mais pas de mouvement de rotation autour de son axe. Cela montre que les traits flagellaires sont extrêmement symétriques à la fois dans le plan ( xy ) et hors plan (z ) directions, qui correspond aux observations en 3D.
Le référentiel d'accompagnement * est un référentiel associé à l'organisme en question à un moment précis. Le corps à l'intérieur de ce système est immobile. Par exemple, un ascenseur en chute libre est un cadre de référence d'accompagnement pour un corps qui y tombe librement, mais la Terre n'est pas un tel système par rapport au corps dans un ascenseur.La projection de la trajectoire du milieu du flagelle (lignes rouges en 2B et 2D ) montre un incroyable éventail de motifs géométriques, des étoiles en rotation aux triangles, carrés et motifs en boucle à symétrie polaire. Le motif flagellaire irrégulier montré en 2D pour les spermatozoïdes flottant sur la lamelle est également observé pour les spermatozoïdes flottant à côté de la lamelle. Cette variabilité des motifs peut être causée par une discordance du décalage de phase entre les composants dans le plan et hors du plan du battement pour chaque cellule, ce qui augmente dans la chronologie du mouvement de roulement. Par conséquent, les motifs flagellaires ne sont pas une caractéristique distinctive des cellules flottant près et loin de la lamelle.
Microscopie et modélisation du flagelle montrant la rotation de la tête et du flagelle autour de l'axe de mouvement.
L'amplitude de l'onde 3D est caractérisée par une courbe enveloppe symétrique en forme de balle, par opposition à la spirale conique (similaire à un tire-bouchon expansible), qui est souvent décrite dans la littérature. Les spermatozoïdes qui flottent loin de la lamelle avaient une forme d'onde plus symétrique que les cellules flottant à côté du verre. Ainsi, la lamelle adjacente est une faible source d'asymétrie pour le battement flagellaire.
Les spermatozoïdes flottant à côté de la lamelle avaient un angle d'attaque préservé * -7 °, avec une orientation moyenne du flagelle du sperme dirigée vers la lamelle.
* — ( ) , .

Image # 3
L'image ci-dessus montre une comparaison des traits flagellaires dans le cadre compagnon (rangée du haut) et le cadre roulant qui l'accompagne (rangée du bas).
La figure 3E montre la vraie nature du battement flagellaire vu du point de vue du sperme, sans aucun mouvement flottant ou roulant.
Le cadre de référence roulant joint ( 3E ) montre que les volets flagellaires sont anisotropes, c'est-à-dire les caractéristiques des ondes dans chaque direction transversale (perpendiculaire au plan de battement), étiquetées « plan b » (plan bleu) et « plan z » (plan rouge), sont nettement différentes.
Si nous comparons les zones gris clair sur les plans bleu et rouge à 3E, on peut voir que le battement est fortement asymétrique dans le plan b et se caractérise par une symétrie gauche-droite brisée ressemblant à une forme en C.
Cette observation contraste fortement avec les motifs symétriques observés dans le référentiel comoving en 3A .
Forme d'onde du flagelle par rapport au cadre de référence fixe du laboratoire (x, y, z).
L'analyse en composantes principales (ACP) d'une forme d'onde tridimensionnelle a permis aux scientifiques de décomposer l'onde flagellaire en plusieurs «modes de forme de base», appelés ici modes PCA. La forme d'onde 3D peut être reconstruite avec une bonne précision avec seulement deux modes de forme d'onde, comme on le voit avec 3B et 3F .
En 3C, on peut voir que les deux premiers modes PCA sont de forme identique, jusqu'à une rotation de 90 °, capturant ainsi la forme en spirale profilée provoquée par le roulement du sperme.
En 3G , cependant, la forme en C asymétrique interne n'est pleinement détectée que par le premier mode PCA. Deuxième mode PCA sur 3G(surligné en orange) introduit de petites déviations perpendiculaires au premier mode PCA (surligné en bleu), indiquant que la forme d'onde peut être décomposée en deux directions d'ondes transversales indépendantes qui sont de nature anisotrope.
L'analyse de Fourier des balayages a permis de reconstituer le mouvement du flagelle en utilisant seulement deux modes de Fourier en 3D et 3H . En bref, chaque signal de swing commun peut être approché par une simple somme de deux fonctions:
f r (s, t) ≈ f 0 (s) + | f 1 (s) | sin (ωt + φ (s))La première fonction f 0 (s) ne dépend pas du temps, elle est appelée "mode statique", et elle fixe l'asymétrie moyenne du signal sur la longueur du ou des arc (s) flagelle (s).
La deuxième fonction est une onde progressive sinusoïdale, appelée «mode dynamique», oscillant à la fréquence captée par le premier pic du spectre de puissance du signal.
L'amplitude et la modulation de phase d'une onde progressive le long du flagelle sont respectivement égales à | f 1 (s) | et φ (s) = arg (f 1 (s)).
Ainsi, la phase porte des informations sur les caractéristiques de l'onde progressive. Par exemple, si la (les) phase (s) ne change pas le long de s, alors le signal n'est pas une onde progressive, mais plutôt une onde stationnaire.
Par conséquent, le mode statique capture toute discordance de forme d'onde. Dans ce cas, la ligne droite noire en 3D reflète la symétrie du balayage dans les plans xy et xz dans le cadre de comoving. En revanche, dans le référentiel roulant ( 3H ) qui l'accompagne , le mode statique (courbe noire) est caractérisé par une grande amplitude asymétrique, rappelant un C. inversé.
Forme d'onde du flagelle par rapport au cadre de référence associé.
Forme d'onde du flagelle par rapport au cadre de référence roulant d'accompagnement.
Le mode dynamique dans le cadre compagnon (courbes rouges) en 3D a une grande amplitude et est très symétrique dans les plans xy et xz en raison du roulement du sperme.
Mais le mode dynamique dans le cadre de référence roulant d'accompagnement (courbes rouges) à 3H a une amplitude de forme d'onde réduite et une direction de déplacement préférée.
La reconstruction de Fourier de la forme d'onde a été réalisée en sommant les modes statique et dynamique (graphes au centre sur 3D et 3H ), ce qui est en bon accord avec les observations d'origine (graphes à droite en 3Det 3H ).
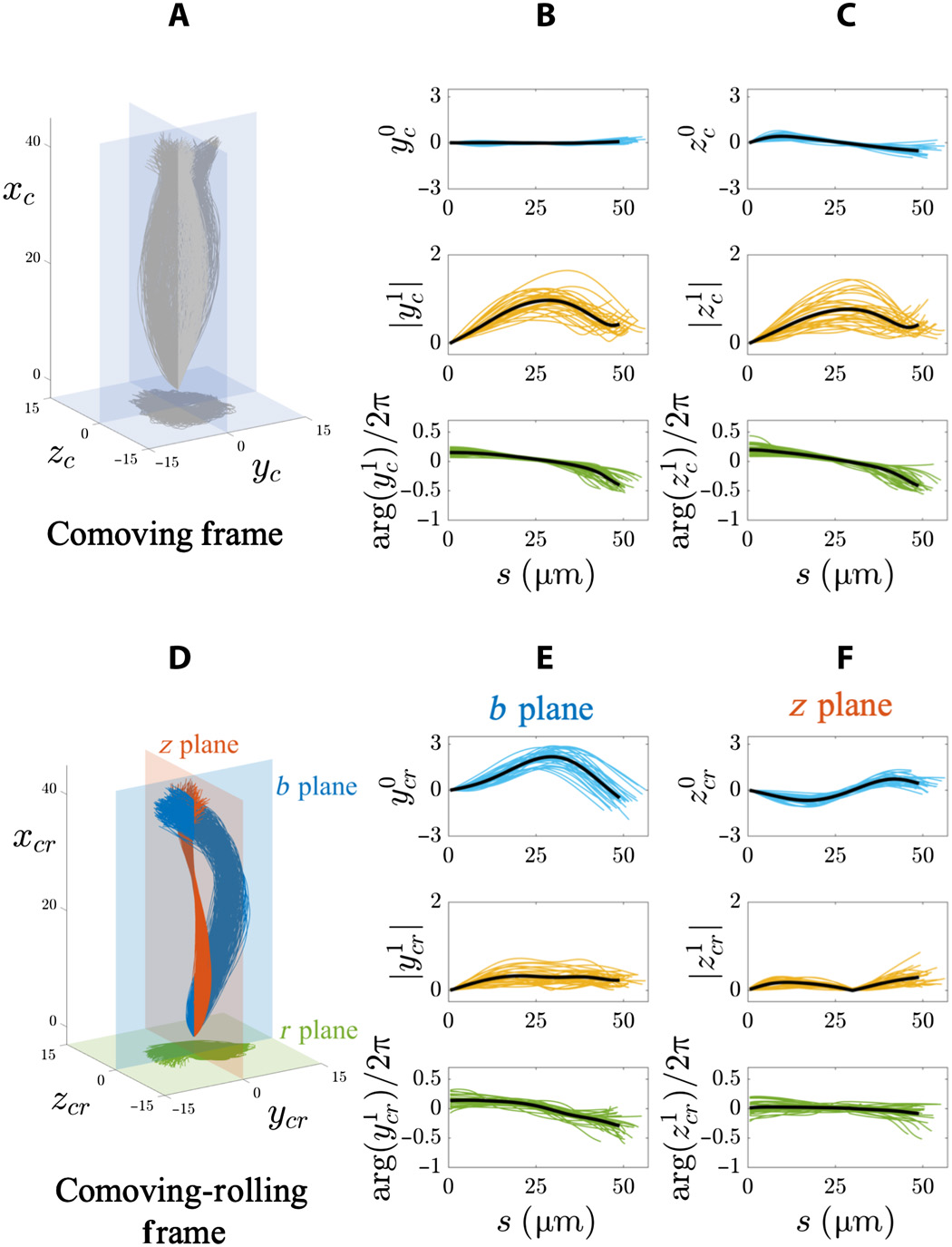
Image # 4 Les
graphiques ci-dessus montrent les résultats d'une analyse de Fourier d'un lambeau flagellaire 3D dans une population de spermatozoïdes flottant librement (20 cellules à côté de la lamelle et 8 cellules à l'écart du verre). Dans le référentiel d'accompagnement ( 4A - 4C ), les amplitudes du mode statique dans les deux sens (y c et z c ) sont très faibles (rangée supérieure des graphiques en 4B et 4C ) en raison de la symétrie des balayages.
De plus, l'amplitude (rangée du milieu) et la phase (rangée du bas) des modes dynamiques (y c et z c ) à 4B et 4Cfixe la symétrie transversale et dépasse l'isotropie dans ce cadre de référence du fait que le spermatozoïde tourne autour de l'axe de mouvement.
Les caractéristiques de l'onde progressive pour les deux coordonnées (y c et z c ) sont les mêmes pour toutes les cellules flottantes: la fréquence est de 4 Hz, la longueur d'onde est de 100 μm et la vitesse de l'onde est de 400 μm / s.
Dans le système de référence roulant d'accompagnement ( 4D - 4F ), des régimes statiques distincts dans les plans b et z montrent que les oscillations ont encore une anisotropie dans une population de cellules flottantes libres ( 4E et 4F ).
Plan b mode statique (y cr) est fortement asymétrique et biaisée vers des valeurs positives (rangée supérieure des graphiques par 4E ). Mais pour le plan z (z cr ), il oscille symétriquement sur la longueur de l'arc de manière sinusoïdale (rangée du haut à 4F ).
L'amplitude du régime dynamique dans le b plan (y 1 cr ) augmente jusqu'à atteindre un plateau (rangée du milieu à 4E ), alors que le régime dynamique dans le z plan (z 1 cr ) est non monotone le long de la longueur d'arc (rangée du milieu à 4F ). Caractéristiques des ondes progressives y cr ( bplan) étaient: fréquence 8 Hz, longueur d'onde 145 μm et vitesse d'onde 1120 μm / s. Caractéristiques d'une onde progressive z cr ( plan z ): fréquence 6 Hz, longueur d'onde 1526 μm et vitesse 5174 μm / s.
De petits changements de phase à de grandes distances sur toute la longueur du flagelle nécessitent des vitesses de propagation des ondes très élevées. Par conséquent, les oscillations du plan z se comportent en fait comme une onde stationnaire pulsant dans le temps.
Microscopie des spermatozoïdes non propulseurs et roulants.
L'ensemble des données ci-dessus suggère que le battement des flagelles utilise deux contrôleurs transversaux anisotropes coactifs, qui ne sont pas trop différents des ondes électromagnétiques en mouvement. Cependant, chaque onde de cisaillement (y cr , z cr ) est la somme des modes statiques et dynamiques: une onde asymétrique progressive le long du plan b (bleu en 4D ) et une onde stationnaire symétrique dans le plan z (rouge en 4D ).
Il est curieux (mais pas surprenant) que la présence d'un verre de protection à proximité de la trajectoire du spermatozoïde ait également son effet sur l'amplitude de propagation des ondes.

Image n ° 5 Le
verre réduit l'amplitude de la propagation des ondes en raison des interactions hydrodynamiques entre le flagelle et la surface dure du verre de protection.
Dans le cadre compagnon ( 5A et 5B ), l'amplitude des deux modes dynamiques (y c , z c ) diminue vers la fin du flagelle du sperme près de la lamelle (graphiques au centre), tandis que les modes statiques restent inchangés (graphiques Haut).
Le mode dynamique z c n'est que légèrement inférieur à y c (courbes bleues) dans les tracés centraux 5A et 5B . Cela contraste avec les profils symétriques et invariables des deux (yc , z c ) modes dynamiques pour les spermatozoïdes détectés loin du verre (courbes rouges dans les graphiques au centre 5A et 5B ).
Si nous prêtons attention au système de référence de roulement qui l'accompagne, alors la nature de l'effet de verre est assez anisotrope, car elle n'affecte qu'un seul plan de roulement, c'est-à-dire sur le plan b ( 5C ).
La lamelle couvre à la fois les modes ycr statiques et dynamiques (graphique au-dessus et au centre à 5 ° C ). Mais le plan z (z cr ) reste inchangé (graphe en haut et centre en 5D ).
La forme du mode statique dans le système de référence de roulement compagnon est la même parmi tous les spermatozoïdes et définit une spirale à droite décentrée indiquée par h (s) et représentée par une courbe noire à 5E . La projection de la spirale sur le plan de roulement (plan vert) est une spirale anti-horaire qui n'a pas de symétrie polaire, c'est-à-dire décalage d'un côté (projection grise à 5E ).
Les spirales de mode statique ressemblent fortement aux spirales logarithmiques que l'on trouve souvent dans la nature. Cependant, dans ce cas, le rayon de la spirale change de manière non monotone, augmentant / diminuant à un rythme plus rapide que celui des spirales logarithmiques ailleurs dans la nature.
Spirale logarithmique droite h (s) à 5Epeut être exprimé en termes de rayon et de hauteur ( 5G ), qui se désintègre de façon exponentielle sur la longueur du flagelle. Tout changement de signe dans h y ou h z fait passer la spirale en rotation dans le sens horaire, créant ainsi une spirale à gauche. Tous les spermatozoïdes flottants ont généré des spirales dans le sens antihoraire. Seuls deux spermatozoïdes avaient des spirales dans le sens des aiguilles d'une montre, bien que de forme identique à 5E . C'étaient ces deux cellules qui ne pouvaient pas nager vers l'avant à cause de certains obstacles sur leur chemin, mais elles pouvaient continuer à tourner autour de leur propre axe de torsion.
Dans tous les cas, la rotation de l'hélice est en corrélation avec le sens du roulement du sperme comme suit: une hélice anti-horaire pour une rotation horaire (vue de l'extrémité arrière) et une hélice horaire pour une rotation antihoraire.
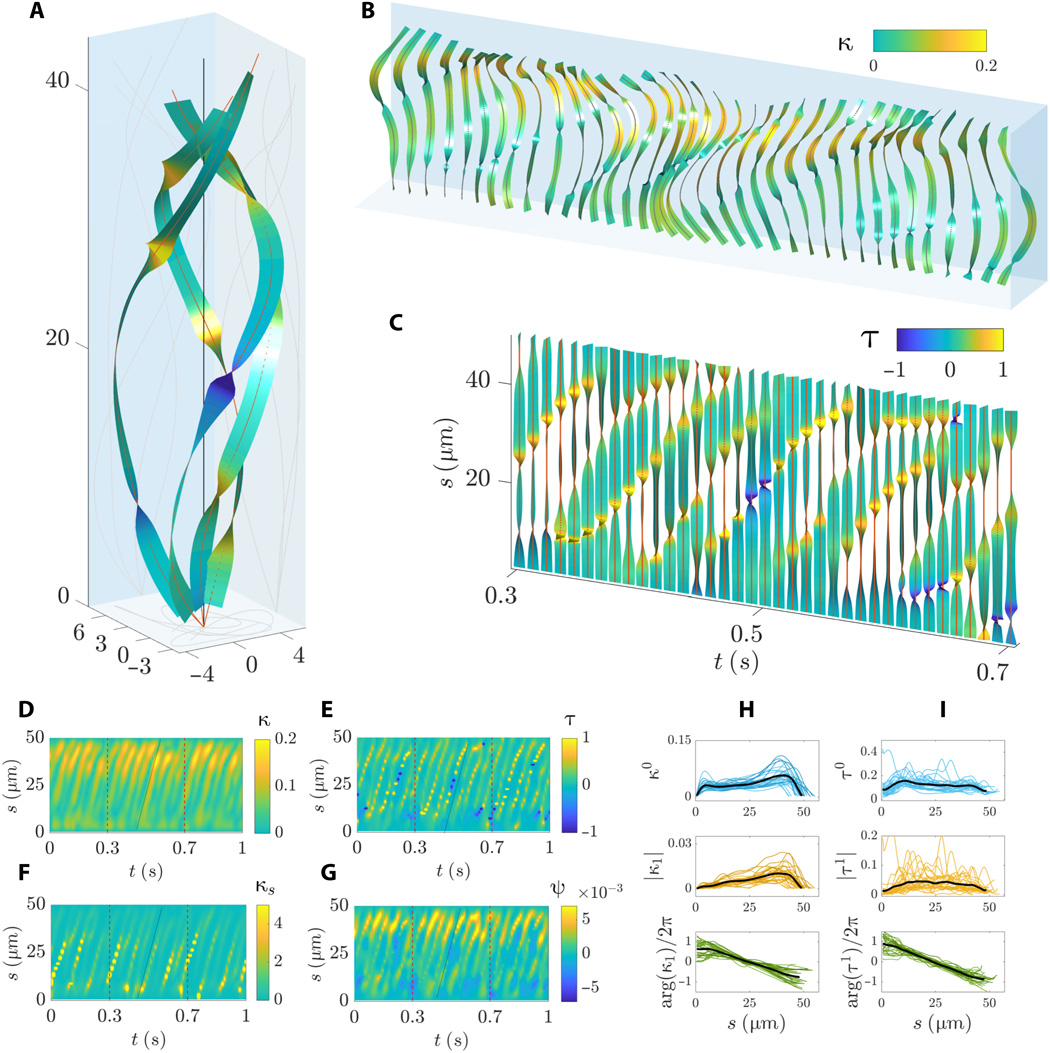
Image # 6 Les
graphiques 6A et 6B montrent une séquence complexe d'ondes progressives lorsque le flagelle tourne autour de son axe de roulement. Les ondes de flexion se propagent linéairement le long du flagelle avec une amplitude non monotone sur la longueur de l'arc, caractérisée par une forte augmentation dans les régions médiane et distale.
La torsion de la forme d'onde est caractérisée par des courbes brusques le long de la longueur de l'arc ( 6B et 6C) avec tours positifs et négatifs simultanés. La forme en spirale de la ligne centrale du flagelle subit un phénomène par lequel des régions de chiralité opposée le long du flagelle coexistent. Cependant, des parties du flagelle avec une chiralité opposée se déplacent pendant le battement ( 6C et 6E ). Les ondes de torsion se propagent à la même vitesse que l'onde de courbure.
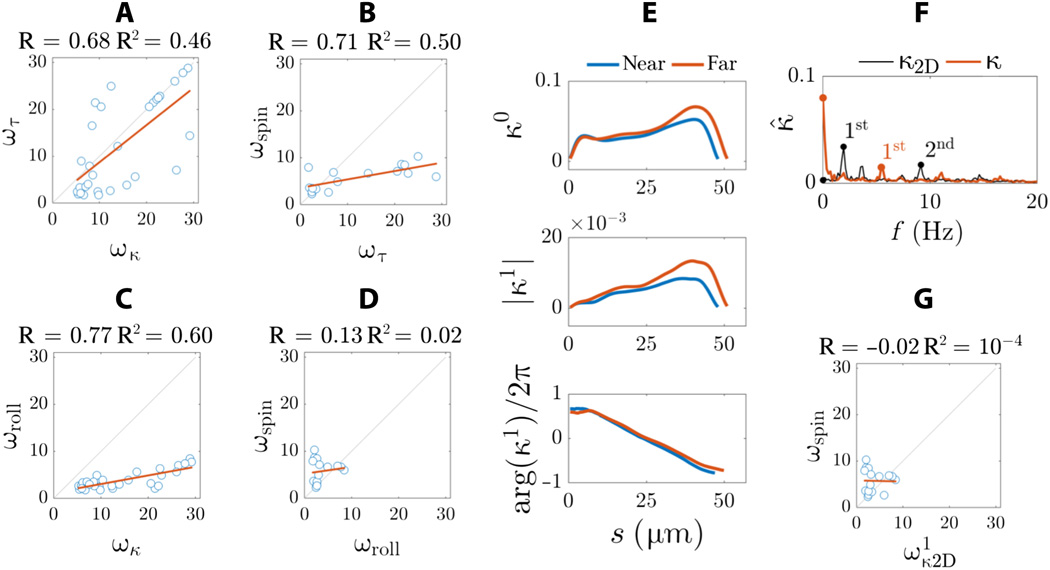
Image # 7
Pour mieux comprendre la différence entre les résultats de microscopie 3D obtenus dans cette étude et les résultats de microscopie 2D qui sont souvent décrits dans la littérature, les scientifiques ont créé une projection 2D à partir d'une forme d'onde 3D.
Le régime statique de courbure bidimensionnelle est très insignifiant ( 7F). En 2D, l'asymétrie intrinsèque de la forme d'onde ne peut pas être détectée. Au lieu de cela, le spectre de fréquence est caractérisé par deux pics de fréquence (marqueurs noirs à 7F ) plutôt qu'un pic de fréquence majeur vu pour une courbure tridimensionnelle (courbe rouge à 7F ).
Pour une connaissance plus détaillée des nuances de l'étude, je vous recommande de consulter le rapport des scientifiques et des éléments supplémentaires .
Épilogue
Dans ce travail, les scientifiques ont pu montrer en pratique qu'en raison de la microscopie tridimensionnelle, il est possible de savoir ce que la microscopie bidimensionnelle ne donnera pas - par exemple, voir les changements de direction de la spirale (changements de chiralité) lors de l'oscillation des flagelles.
À première vue, il peut sembler que les spermatozoïdes flottent simplement vers l'avant, remuant la queue. Cependant, un examen détaillé de la cinématique de ces cellules a montré que ce processus est beaucoup plus compliqué. Le battement et le balancement des flagelles, la rotation de la cellule elle-même - tout cela permet au sperme d'avancer.
Certaines de ces informations étaient auparavant disponibles, mais tous les détails n'ont pas été détaillés en raison des limites de la microscopie 2D. Comprendre comment les spermatozoïdes se déplacent peut beaucoup aider dans le domaine de la médecine de la reproduction, disent les scientifiques. En particulier, les nouvelles données élargiront la gamme de paramètres par lesquels les cellules sexuelles malsaines des hommes sont déterminées, ce qui est la cause de l'infertilité chez environ la moitié des couples.
Dans tous les cas, mieux nous comprenons certains phénomènes et processus, en particulier ceux qui se déroulent dans notre propre corps, plus il est probable que nous améliorons la qualité de notre vie.
Merci pour votre attention, restez curieux et passez une bonne semaine de travail, les gars. :)
Un peu de publicité
Merci de rester avec nous. Aimez-vous nos articles? Vous voulez voir du contenu plus intéressant? Soutenez-nous en passant une commande ou en recommandant à des amis, le cloud VPS pour les développeurs à partir de 4,99 $ , un analogue unique des serveurs d'entrée de gamme que nous avons inventé pour vous: Toute la vérité sur VPS (KVM) E5-2697 v3 (6 cœurs) 10 Go DDR4 480 Go SSD 1 Gbps à partir de 19 $ ou comment diviser correctement le serveur? (options disponibles avec RAID1 et RAID10, jusqu'à 24 cœurs et jusqu'à 40 Go de DDR4).
Le Dell R730xd 2x est-il moins cher dans le centre de données Equinix Tier IV à Amsterdam? Seulement, nous avons 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2,6 GHz 14C 64 Go DDR4 4x960 Go SSD 1 Gbps 100 TV à partir de 199 $ aux Pays-Bas!Dell R420 - 2 x E5-2430 2,2 GHz 6C 128 Go DDR3 2 x 960 Go SSD 1 Gbps 100 To - À partir de 99 $! Lisez à propos de Comment construire l'infrastructure de bldg. classe avec les serveurs Dell R730xd E5-2650 v4 au coût de 9000 euros pour un centime?